There are awesome news about one of the most unusual cetaceans in modern history, an enigmatic hybrid between a narwhal (Monodon monoceros) and a beluga whale (Delphinapterus leucas).
The specimen was shot at Disko Bay, Greenland, in 1986 or 1987 and the skull – the only remaining relic – collected there in 1990 and sent to Copenhagen. I´ve been fascinated by this freak of nature since I´ve read about it for the first time in 2007. From all the photos available at that time I tried to make a reconstruction of it, but I was not really happy with it. In 2017 I had finally the chance to examine the original skull of this specimen in the archives of the Zoological Museum at Copenhagen, what gave me some highly worthy insights into its extremely weird anatomy. Besides the skull only a few vague descriptions of the living animal were known, so dealing with it was like dealing with the remains of a prehistoric animal only known from fossils. This made it of course even more interesting to make a reconstruction of the living whale, which was subsequently termed „narluga“ by some people.
I want to thank here Eline Lorenzen and especially Daniel Klingberg-Johansson for the chance to visit the non-public archive where the skull is located. The public exhibition of the Museum has many incredible and absolutely priceless specimens (more about some of them in a future blogpost), but as usual a lot of the really good stuff is in the archives. Thanks again Daniel for this awesome tour to the museal catacombs!
In this blogpost I want to give you some more background information which is known about the narluga, and how I tried to give the skull a face and a body. As I already wrote it was shot in 1986 or 1987, by an inuit hunter named Jens Larsen. The correct date is not known, as Larsen who didn´t remember the year for sure. It was discovered by Mads Peter Heide-Jørgensen on the roof of a toolshed at Kitsissuarsuit, which is located at the mouth of Disko Bay, West Greenland. Larsen had perceived it unusual enough to save the skull and donated it for further examination. He sadly could not give much more information about it, unless that it had flippers which resembled those of a beluga and a fluke more similar to those of a narwhal, but a coloration which was not white like a beluga or white and dark-gray mottled like a beluga but uniformlys gray and unlike those of those two species. What was even more interesting was the fact that he remembered that there were two additional specimens, which were shot as well. One was lost during the hunt (modern inuit shot belugas and narwhals usually with guns, so a lot of them just sink to the bottom of the sea and are lost), the head of the other one was saved and left near the water for maceration, but never retrieved.
Articulated skull of the Narluga, Mikkel H. Post, Natural History Museum of Denmark
After the skull was sent to Copenhagen, it was examined and compared to various skulls of belugas and narwhals from the collection of the Zoological Museum. It exhibited several unusual features. It was somewhat larger and more elongated than the means of the skulls used for comparison. Its sutures of the premaxilla/maxillare were fused, what indicated that the specimen was already comparably old. The most bizarre feature of it was however the dentition. It was totally unlike any other known whale.
Narluga maxillary dentition, photo Markus Bühler
It had originally ten upper and eight lower teeth, from which one of the maxillary teeth was already lost when the skull was found. In the living narluga, six or seven of the upper teeth were likely exposed inside the cavity of the mouth, as they showed also signs of abrasion. The three most anterior teeth however apparantly never erupted through the gums and remained mainly within the bone. Their appearance was extremely strange, very thin, long and horizontally orientated within the bone, a bit similar to the unerupted vestigal teeth of narwhals but lacking their typical knog at the apical end. The more distal teeth were shorter and more curved. According to Lars Jensen, the specimen which sank and was lost had even some visible protruding upper teeth.
Extracted teeth, photo Eline Lorenzen, Natural History Museum of Denmark
The mandibular teeth in contrast were all eruted, very large and massive, assymetrical in shape and orientation. In the living animal they were likely visible even when the narluga had its mouth closed. Two of those teeth show unusual curving and slight twisting which must have occured during growth, and even longitudinal grooves vaguely similar to those seen in erupted narwhal tusks.
Right mandibular teeth, photo Markus Bühler
The most distal tooth on the left side was very small and much more posteriorly located then the fourth tooth on the right side. It was sadly lost, but the orientation of the tooth socket indicates quite clearly that it was pointing backwards.
Narluga skull, photo Markus Bühler
At the time of its first examination it was still not known if it really was a hybrid between a narwhal and a beluga, or just a really unusual narwhal with extreme dental anomalies. Genetic examinations however clearly showed that it really was a male hybrid, with a narwhal mother and a beluga father. Another really interesting result from the examination of carbon and nitrogen from the bone collagen revealed that the narluga differed in diet from both belugas and narwhals. The fish on which it preyed were apparantly from more benthic regions than those usually consumed by narwhals and belugas. Perhaps this diet resulted from the unique dentition of this individual, which made it harder to hunt and catch the more common prey animals of its parental species.
When I worked on the reconstruction of the living animal, I tried to take everything known about it into account. The shape of the skull, the orientation and abrasions of the teeth and the description of the external features like flippers, fluke and coloration, as well as the anatomy and life appearance of Monodon and Delphinapterus. I gave the narluga the head-to-body-ratio of a beluga and tried to make its external body shape intermediate between those of a narwhale and a beluga. Narwhals have streamlined bodies, but belugas have a very distinct neck area and weird fat deposits at their belly area.
Narluga reconstruction
By far the most difficult part of the reconstruction was the head and mouth. Narwhals and belugas differ in several aspects of their cranial soft tissue. Belugas have – quite unusual for extant cetaceans – quite fleshy and flexible lips, which can be moved into a lot of shapes. Even more problematic was their ability to change the shape of their melon area, because this made it really hard to find a good reference with which I was happy, since every beluga photo looks different – even if it´s the same indivudual but with different melon shapes. I made many tries until I found something with which I was mainly happy and still somewhat intermediate between narwhals and belugas. I didn´t gave it beluga lips, but made them at least a big more pronounced than those of narwhals.
Narluga head reconstruction (still without the fourth left mandibular tooth)
The upper teeth are not visible here (they usually never are visible from lateral views in most odontocetes). I directly used the original shape of the lower teeth of the articulated skull from an earlier press release to get them as correct as possible and to show also such features like the partial torquing and spiraling. The fourth left tooth was missing, so I had to reconstruct it from the position and size of its tooth socket, which show that it was much smaller than the other mandibular teeth, assymetrically distally located and backwards-pointing.
Reconstruction of the missing fourth left mandibular tooth. You can see the very strong assymetrical location
I nearly missed to portray a minor but important detail until the very end, the blowhole which is nearly invisible as this position. But luckily I just remebered to draw it before I sent the final version to Eline Lorenzen.
The coloration was described as gray and unlike those of narwhals or belugas. This is not that unusual that hybrids don´t just show intermediate patterns or colors of their parental species but unique new traits. Because both belugas and narwhals are comparably uniformly gray at young age, I used them as reference for the color of the narluga, with a very slight mottling reminiscent to those of juvenile narwhals. Because it was an older specimen, I gave it a few scars, because it is very likely that it received some from rocks and ice during its life.
To make the scenery a little bit more atmospheric I added a small swarm of polar cods (Boreogadus saida) in the background. This species occurs in the area where the narluga was shot, from river mouths to depths up to 900 m, what makes it a very well fitting faunal element, especially as it´s a very common food item for both belugas and narwhals.
Polar cods (Boreogadus saida)
I made the flippers mainly based on the large pectoral fins of belugas, but somewhat lesser developed, but slightly darker than the rest of the body as seen in narwhals. The fluke shows the unusual „backwards“ shape of male narwhals (those of females look more normal), just somewhat lesser expressed. Like all my other digital art projects (for example my recent Basilosaurs), this was made with GIMP. The whole project took several months until it was finally finished, but it was also a really interesting and exciting project. A lot of it was surely speculative, even if based on some known known facts. Perhaps anytime we will see another, perhaps even living narluga, which will show how close my rendition was to the true life appearance.
Sources:
Heide-Jørgensen, M. P. & Reeves, R. R. Description of an anomalous Monodontid skull from west Greenland: A possible hybrid? Mar. Mamm. Sci.9, 258–268 (1993).
Mikkel Skovrind et al. Hybridization between two high Arctic cetaceans confirmed by genomic analysis, Scientific Reports (2019).
Last year I was involved into a fantastic project, the production of the wildlife documentary „Into the Forest: Reptiles & Amphibians“ by my good friend Bryan Maltais.He made already several other shorter documentaries, including the award-winning production „Metamorphosis“ about the amphibian life in a wetland in Colorado. But this was by far his biggest project. I want to write here about the filming and the background of „Into the Forest“, which is now also available on Amazon Prime, both in English and German.
Bryan spent his early youth with his German mother and American father in Germany but later grew up in the U.S. Over the decades he regularly came back to visit his second home and the German part of his family. During those vacations he regularly visited a forest near Denkendorf, a village not far away from Stuttgart, in the southwest of Germany. It was this forest where he came for the first time into contact with the European herpetofauna and other native animals. He later studied wildlife biology and developed a particular interest into reptiles and amphibians. Over the years Bryan became a professional nature and landscape photographer and produced several documentaries, mainly about reptiles and amphibians.
Bryan Maltais at Mills Lake inside Rocky Mountain National Park
We met for the first time in 2014 after I discovered his documentaries on YouTube and realized that Denkendorf is in fact comparably close to my own place of residence. Over the years we met for several more times for herping tours and I introduced Bryan to various other interesting locations like the habitat of my local green lizard (Lacerta viridis) population or the Black Forest with its melanistic European adders (Vipera berus).
Bryan taking close-up photos of a green lizard
But „Into the Forest“ became a much bigger and much more extensive project than just a few tours to the woods. Bryan spent three whole months in Germany to document the rise of nature from the last snow of march to the early summer in june. He was filming for nearly every single day and sometimes even during night, and when he wasn’t filming he worked on editing and cutting the videos.
To produce a high quality documentary he used a 4k camera so you can also enjoy it on a big screen in TV quality. He made slow motion and fast motion videos, underwater-and night vision footage and made also some amazing landscape videos with a drone (which had tragically a fatal accident, luckily all the important drone footage was already filmed at this time).
View over the Black Forest. Screenshot from „Into the Forest“
During this three months we met as often as I could to show Bryan several more interesting locations like a pond which is especially well suited to observe mating common toads (Bufo bufo). One particular experience was a tour during a rainy night to a small valley just quite close to where I live. This area is a very good location for one of the most charismatic European amphibians, the flamboyant fire salamander Salamandra salamandra. Admittedly I was never there after dusk, and our tour topped easily everything I’ve ever seen during a salamander herping tour. We saw so many fire salamanders that we couldn’t even count all of them, but all in all we’ve seen about 60 specimens. In this area the populations of the striped fire salamander Salamandra salamandra terrestris and the spotted fire salamander Salamandra salamandra salamandra overlap and both forms also interbred. As a result of this we encountered an enormous range of different patterns. We also found a specimen which just caught an earthworm, a pair which was mating and best of all several females just giving birth to their larvae.
Filming fire salamanders during a rainy night is not exactly the most comfortable thing you can imagine
The fire salamander also plays a key role in the documentary, but not just for its charming appearance but also to raise awareness about the dangers of the chytrid fungus Bsal which entered the European continent from Asia over the amphibian pet trade. Bsal already eradicated whole fire salamander populations in Belgium and the Netherlands, and has already reached northern Germany. Sadly, it is likely to continue spreading even further through Europe.
Female fire salamander giving birth to its larvae. Photo by Markus Bühler
Another nightly tour brought us to a local population of European tree frogs (Hyla arborea). The loudness of their croaking was just incredible. This was for me also the very first time to see European tree frogs, as this species has sadly become extremely rare, mainly as a result of a lack of fitting habitats and small periodically draining ponds where they can successfully spawn. I was also quite surprised to learn about this population which is also quite closely located to some of the other areas where Bryan filmed.
European tree frog, photo by Markus Bühler
Of course the wonderfully exotic-looking (and in this case allochthonous) green lizards (Lacerta bilineata – or is it viridis? Things are complex here) of the local population here around were included as well. Bryan could even film a mated pair, and the drone footage provided some great views of their habitats within the centuries old vineyards.
Green lizard
Another reptile we hoped to find remained however elusive. The year before we made a trip to the Black Forest (which is also only about an hour to drive from here) and were able to find two specimens of melanistic European adders.
Melanistic form of the European adder, the „hell adder“
But this year we had sadly lesser luck. During our first trip to the Schliffkopf, one of the highest elevations of the Black Forest, it was despite the early time of spring extremely hot, well over 25 Celsius. On some places there was still half a meter of snow, but the heat of the sun was just too much for the adders.
Not a tundra area at Scandinavia but an upland plain at the Schliffkopf. Photo by Markus Bühler
At least we could observe some particularly nicely colored European grass frogs (Rana temporaria) in the act of mating, and this sequence was also included into the final documentary.
Mating grass frogs. Photo by Markus Bühler
We made later two further trips, but every time the weather conditions were against us. At the second tour to the Schliffkopf mountain it was quite the opposite of the last time, it was very cold with a lot of wind and fog, but next to no sunshine. Again pretty bad conditions for adders. However the fog over the forest provided some spectacular landscape shots.
Fog and clouds over the Black Forest. Photo by Markus Bühler
The view from the slopes around the Schliffkopf is really great, you can even see the Alsace behind the river Rhine.
Bryan looking to France
Our last tour to the Black Forest lead us to Kaltenbronn, an area with several bogs where we found the two adders the year before. Sadly all we found that time was a single freshly road-killed adder.
Bog area at Kaltenbronn. Photo by Markus Bühler
A photo of me from our more successful adder-tour in 2017:
One highlight was also a herping excursion to Croatia. There would be so much to say about the many things which we have seen there, but this would be way too much here. We made a tour from the island of Krk to to the very southwest of the Croatian mainland, down over Istria to the island of Cres and again back to Krk. We saw various interesting reptiles, birds and a lot of really unusual invertebrates and spectacular landscapes. We didn’t find as many reptile species as we hoped, especially snakes and scheltopusiks (Pseudopus apodus) proved to be quite elusive and hard to find. I spied only a few of scheltopusiks at the first day at Krk, but it was not possible to take any photos. One species for which we had especially big hopes to see it was the horned adder (Vipera ammodytes), one of the most venomous snakes of the European continent.
Horned adder, Krk.
It was still a lot if luck involved, but during our systematic search I could detect a tiny horned adder between some stones. This enigmatic little snake was surely one if our greatest finds during the excursion.
Bryan and the horned adder, island of Krk, Croatia.
To have at least a more southern cousin of the European adder on screen, it was also included within a short segment about our excursion. There would be still so much more to say, but I will make a break here, in hope to write anytime a future blogspot about our tour to Croatia. You can see some more photos on Bryan’s website.
A considerable part of the documentary was filmed in the Schönbuch forest, a large forest area near Tübingen. The deep and wide-ranging woods provided various interesting locations for filming, from game enclosures with red deers and wild boars to large ponds full of tadpoles. Germany is quite densely populated and even most of the nature reserves are in one way or another altered by human activities. In many cases this already happened centuries ago, so the landmarks of human activities are often deeply interwoven within the surrounding landscape.
One of the big ponds of the Schönbuch forest. Photo by Markus Bühler
One especially nice remnant of human culture was also included, the old monastery of Bebenhausen which is located just in front of the Schönbuch forest. Surrounded by centuries old wooden framework houses, its gothic architecture, ancient walls and old towers made it also a perfect location for drone footage to show the full range of the complex.The monastery was founded in 1190 and after the reformation it was among others used as a hunting lodge and the late residence of the last king of Baden-Württemberg in the early 20th century.
The old monastery of Bebenhausen. Photo by Markus Bühler
We visited also the Alter Aichwald („old oak forest“) near Denkendorf where Bryan made much of the filming of fire salamanders and other featured amphibians. We were there already years ago on our first fire salamander tour.
Fire salamander at Alter Aichwald
Bryan showed me the big badger den which he discovered and where he could make some great footage with a game camera.
I know many of those locations for nearly my whole life and have countless wonderful memories about them. To see them now in high quality documentary feels nearly somewhat surreal, especially as I was on site during many of filming sequences. I learned a lot of new things and made many new experiences, and it makes me really proud that I could contribute a little bit to this wonderful documentary. Bryan did really an amazing job to produce the whole documentary himself. He did all the cutting of the scenes, the selection and arrangement of the background music, the script (which I translated and adapted for the German version, entitled “Der magische Wald: Reptilien und Amphibien) and the digital enhancement of certain scenes. Normally such documentaries are made by whole production teams or even specialized companies.
Screenshot from „Into the Forest“
„Into the Forest“ became a really wonderful documentary especially as it focuses on animals which get despite their fascinating lives usually only little media coverage. I think it will be also quite interesting to see for everyone interested in animals, no matter if you’re European and familiar with some of the featured animals or if you are living anywhere else on the world. I could imagine that for non-European watchers it could be especially interesting to learn about some of the enigmatic animals which still dwell in such a non-exotic area, and also about the surrounding landscapes and history of Southern Germany.
You can read more about the documentary at the official website Into the Forest.
Bryan has also many of his nature and animal photos on MacroPhotoBug, wildernessshots and his instagram and Facebook-page. His documentaries are available on his YouTube channel.
Many thanks also to all who wrote suggestions, ideas and constructive criticism during the editing of “Into the forest”.
If you read a comic or watch a cartoon which includes any crocodiles or alligators, you will notice that they are nearly universally green. The same applies to crocodiles in books for children, on logos or sometimes even drawings in nature books. But are crocodylians really green? To answer this question, it is important to take a look at the various modern crocodylians which includes the crocodiles, the alligators the gharials.
If you are only familiar with „normal“ crocodylians like Mississippi alligators, nile crocodiles or saltwater crocodiles, you will be possibly surprised by the diversity of colors and patterns, especially if you also take a look at the juveniles which are often much more contrast-rich than older specimens.We find near white and full black, a big range of grey, but also many shades of yellow and brown which can be even reddish. And of course there are combinations and mixes of all of them as well as fully black patterns.
Siamese crocodiles (Crocodylus siamensis) are quite flamboyant for crocodylian conditions. Wikimedia Commons, photo by Marco Almbauer
But what about green? This is not that easy to answer. We clearly see not a single crocodylian as green as the eponymous green lizard or the green iguana. At best we find something that falls somewhere in the range of greenish, for example in Nile crocodiles. But even here it seems to be not a real propper green but more a mix between yellow and grey that appears green.
Nile crocodile, Hagenbeck Zoo Hamburg
It’s quite interesting that even ancient Egyptian artists already sometimes exaggerated this to a much stronger green.
Fragment of a sarcophagus of Peteamun, 23.-25. dynasty. Museum Alte Kulturen, Schloss Hohentübingen.
The Nile crocodile Crocodylus niloticus and the West African crocodile Crocodylus suchus were also the first crocodylians which became known to European cultures. The Romans even imported some crocodiles from North Africa to exhibit them or to slaughter them in gladiator fights. We still even find some surprisingly realistic artistic depictions of crocodiles from Roman times, which were quite likely made by people who had actually either seen then themselves or had at least very good references. But besides those we find also some much more grotesque depictions as well.
West African crocodile (Crocodylus suchus), Copenhagen Zoo. Wikimedia Commons, photo by Lars K. Jensen
It’s
not really that surprising that later medieval depictions of
crocodylians were usually just vaguely naturalistic at best. But the
green color was still a comparably common feature. During the
renaissance, when cabinets of curiosities became chic, taxidermy
crocodylians were quite popular items for such collections. Trading
connections and the discovery of the New World and other exotic areas
lead at that time to a tremendous increase of new animal discoveries
and a much better understanding of the natural world. The imported
taxidermy specimens of crocodylians were surely not green what makes
it even stranger that the stereotype of the green crocodiles managed
to persist in art.
If we look at other species we find some other crocodylians besides C. niloticus which have a vague greenish appearance. But even if we keep the slight ability of color change of some species in mind, we still don’t see anything even remotely close to a real green. The only green crocodylians which we find in the real world have their coloration from a layer of algae.
Algae-stained nile crocodile from crocodile farm Djerba Explore, Wikimedia commons. Photo by Ad Meskens
It’s hard to say how much such algae-covered crocodiles, caimans and alligators contributed to the stereotype. In some cases it’s not even that obvious anymore that the original coloration is altered by a fine layer of tiny water plants and even species which have normally not even a vaguely green touch can appear to be of that color.
Algae-stained dwarf crocodile (Osteolaemus tetraspis), Tierpark Berlin. Normally this small crocs are of a dark blackish grey.
I could imagine that this really was a factor which contributed to the cliché of the green crocodiles. Many early travelers and explores could probably see crocodylians only from some distance and usually in the water.
Orinoco crocodile Cocodrilus Intermedius – also showing the dirty mix between yellowish and grey which appears vaguely greenish. Wikimedia Commons, photo by Mauriciogq
To
put the initial question – are crocodiles really green – in a
nutshell, I would say that some are perhaps a little bit.
False gharial (Tomistoma schlegelii) with light-brown main color. Wikimedia commons, photo by Junkyardsparkle
But yet they are still far away from the common stereotypes. That makes it even harder to understand that we still see reconstructions of prehistoric crocodylians with truly comicesque green colors. I am fully aware that we cannot rule out the possibility that prehistoric crocodylians could have had a wider range of colors, especially as they filled many more ecological niches than the remaining modern lineages. But makes it really sense to reconstruct those forms with a color we see in no extant crocodylian species? I really don’t think so. I have to emphasize here that I am refering here only to amphibious crocodylians and crocodyliformes, and not the various terrestrial forms which had totally different ecologies. On a sidenote, I am also not refering to marine forms, which had probably again quite different color schemes and patterns, but I also really doubt that any of them was green either.
The nearly fully blackish coloration of the Mississipi alligator is quite popular among modern crocodylians. Photo taken at Wilhelma Zoo Stuttgart (but the alligators are no more on display today).
The range of colors we actually see in modern crocodylians gives paleoartists already so many possibilities to create interesting new color schemes and patterns, that it’s not even necessary to include green at all. I wrote in an earlier blogpost about a certain tendency to depict marine reptiles with extremely elaborate color scheme and why this is somewhat problematic and contrary to what we see in most bigger marine vertebrates.
Juvenile mississippi alligators have striking patterns, quite unlike the adults. That´s something really important to keep in mind when depicting juvenile prehistoric reptiles. Wikimedia commons, Photo by AThiele.
Mark Witton gave recently a really interesting and thought-provoking talk about monsterization in paleoart. This included also the importance to depict prehistoric animals not like semi-fantastic beasts but as real animals to enable people to really understand how this were once living organisms and not concept art for a sciene fiction movie.
Juvenile black Caiman from Moscow Zoo. The pattern and the strong contrast between jet-black and nearly fully white and creme-yellow is really awesome. Wikimedia Commons, photo by Rigelus
Don’t get me wrong, I really don’t say that we should now depict anything in grey, brown or yellowish colors like all the dinosaurs in the first Jurassic Park movie (except the imaginary frill of the Dilophosaurus of course…). But we should – even if we create new patterns and color schemes – still not forget to see how real animals are looking in life. In the case of extinct crocodylians we even have the chance to see some of their relatives still alive – something impossible for many other prehistoric animals without any extant relatives.
Cuban crocodiles have wonderful contrast-rich patterns even as adults. Wikimedia Commons, photo by Trisha M Shears
So, does it make sense to reconstruct extinct crocodylians and crocodiliformes with colors we don’t find in any extant species? Not really. Even more so when it comes to prehistoric crocodylians which actually are still comparably closely related with modern forms and share even a similar ecology.
A quite demonic-looking Cuvier’s Dwarf Caiman (Paleosuchus palpebrosus) in Paignton Zoo, Devon, UK. Wikimedia Commons, Photo by Nilfanion
That’s
why I would not reconstruct an animal like Purussaurus, Mourasuchus
or any other amphibious crocodylian or crocodiliform with a green
coloration which is more pronounced than the dirty greyish-yellow of
Nile crocodiles. I really think there is a reason that no modern
crocodile, alligator, caiman or gharial is really green, no matter if
they hunt fish, turtles or zebras and no matter if they live in a
jungle river, a swamp area like the everglades or a savannah habitat.
I was recently able to acquire two fresh partial ostrich legs from a breeder. This was a perfect chance to study the external and internal anatomy of this giant bird’s legs and feet. I could take a really close look at things that I could neither see in a living ostrich in a zoo nor in a mounted taxidermy specimen. For me this was particularly interesting because I gould get firsthand data about anatomical details for sculptures and illustrations for which it’s very hard to find good references.
The feet of the ostrich are weird, even grotesque compared with normal bird feet. They are quite likely the most derived feet of any extant flightless bird. Functionally they are closer to the feet of an ungulate than to those of a typical avian. The first toe is enormous in size and has a wide and nearly hoof-like nail whereas the second toe is much shorter, flattened and has no more claw at all. Instead it has a very tough and robust skin with soft pads at the proximal bones. Those pads are made of rounded fat deposits which are embedded within the surrounding connective tissue. The second toe reminded me a lot on the soft-padded toes of camels. The dissection and removal of the skin from this toe without a lateral cut was just awful, and I am not willed to do this ever again.
If you ever wanted to know how an ostrich legs looks like witout skin – here it is. At this stage the soft tissue had already dried and shrunken and the massive sinews had lost a lot of their original thickness.
Before you continue reading I have to apologize for not using that many references and super-correct descriptions for the underlying anatomy of the foot and leg. I really wanted to focus particularly on the scales, and the very large number of photos here made it already quite time-consuming to write the blog post. As there was also a lot of discussion about this topic, I also hoped to finish it today. If I find some more time, I´ll try to make an edited version with some more background information. I also realized when I wrote this that I had lesser usable photos than I thought, because the quality of some of the photos I took was not good enough.
What I found particularly interesting was the large variation of different scale types on different parts of the leg. Of course this is especially good to see in an ostrich, simply because it is such a huge bird. I think that even many people who are familiar with birds are not really that aware of the complex patterns and differences of the scales on bird legs. I write this all also not only to tell you something about the anatomy of modern birds but also as a help for paleoartists to provide some references which could be helpful for depictions and sculpted reconstructions of dinosaurs and prehistoric birds. Luckily we have several really good fossils and also some very nice track fossils which show us partial skin patterns of certain dinosaurs. I was quite surprised to see nearly the whole range of all known dinosaur scale variants on this single ostrich foot and lower leg, plus some more which I’ve never seen in a fossil.I thought it would be best to start at the base of the legs and go further down up to the toe tips.
There was an area of extremely thick skin at the ankle area. The scales were highly keratinized and formed some kind of knee pad to protect the underlying parts.
You can also see how a lot of dirt was pressed in-between the scales of the ankle area.
This is the ankle area in side view. You can see the „knee pad“ area and the highy wrinkled and much softer and thin skin behind it.
This photo shows the scales at about the midle of the leg. If you take a close look you can already see how the scales differ at the front side and back side.
Some more photos from the middle of the leg:
I found this scales especially interesting, as they have quite well pronounced bumps.
Those lower legs consists nearly only of bone and giant sinews wrapped in skin.
You see here the area where large irregular and somewhat thickened scales are forming into the pronounced scutes.
On this photo you can see the large scutes which are located in the area above the foot. At this little segment you can already see a whole lot of different sizes and shapes of scales.
The skin at the backside of the tarsometatarsus is very soft and loose and forms very big folds to allow enough flexibility.
Here´s again another view of the whole foot.
The big scutes don´t form a line up to the toe tip but are interrupted by smaller scales…
…just to form again large scales over the big toe. You can again see an interruption of the scutes where they break into a double line of smaller scutes.
The largest scutes are at the proximal end of the toe and form big shields.
Now the smaller lateral too is really weird. You can see thick and polygonal scales with highly keratinized surfaces on the left which are similar to those at the ankle area. But also their transformation into the large scutes on the upper sides of the toe and rounded, smooth scales with large areas of skin in-between on the sides.
Another detail of the photo above.
Now let´s take a look at the bottom of the foot. The nail is very short, wide and blunt and feels nearly like a hoof.
Most of the scales on the sole of the toes are very small and highly keratinized and form dense pads.
But what the heck is going on here? The scales at the centre of the foot form into bizarre upwards turned structures.
Now this looks really super-gnarly. Can you imagine how the feet of a large theropod or even sauropod could have had similar or even more extreme scale formations?
The area behind this spikey pads is again formed from very small, very irregular scales.
This really large number of very different and highly specialized scales is an important reminder that even fossilized patches of dinosaur skin show us still only a small fraction of their bodies. We also have to keep in mind that this ostrich leg is just an ostrich leg and should not be used as a universal reference. The sizes, patterns, shapes and functions of the scales on bird legs are much more diverse than many people think and we can be next to sure that there were also a lot of different scale types present in dinosaurs, even if they were usually not as elaborate and dragonesque as in some modern squamates. As this topic evoked a lot of interest in recent online discussions about dinosaur skin I decided to write some more blog posts dedicated to this, especially to feature some more non-standard scale patterns on bird legs.
Basilosaurus was just big in the news when a new study about its feeding habits and the predation on young Dorudons was published. This is probably a good time to feature a new reconstruction of the „king lizard“ that I just finished. Originally this was not even made to jump on the Basilosaurus bandwagon but I was looking for an animal which I could use for a certain little art project, and it just fitted perfectly, mainly due to its extreme body proportions. But it became quickly more and more complex and the initial little illustration evolved into much more. I wanted to make a reconstruction which included the aspects which I already discussed in length in my earlier blogposts about the cranial soft tissue of archaeocetes and the highly likely presence of facial vibrissae in this animals. Now I want to explain in detail how and why I reconstructed my Basilosaurus in the way I did.
The Body
Basilosaurus full body reconstruction with updated number of vertebrae
I used the famous skeletal drawing of Basilosaurus isis by Kellog (1936) as main reference, however not the original but a modified version by Cameron McCormick with the updated number of vertebrae based on the examination on the vertebrea counting of three overlapping skeletons (Gingerich et al 1990) and a paper by Zalmout et al. 2000.
Basilosaurus by Kellog (1936), modified with additional vertebrae by Cameron McCormick
As a result of this, this Basilosaurus became even more elongated than Kellogg’s original drawing indicated. This resulted also in some problems, as the whole body was so strongly elongated that I could hardly work on it on the screen at once. Instead I had to work on sections and resize the view again and again to see how it works. One of the main references for its body shape was the ribcage, which shows us how thick its body was about. As Basilosaurus had an extraordinarily long abdomen, I made it somewhat more voluminous than the chest, similar to what we see in many modern cetaceans. To find the basic shape of the body was the most important thing to start until I went more into detail. I had to change my initial draft considerably when I re-checked the ribcage and forelimbs with the skeleton to get the position if the shoulder girdle in the correct way.
Head and mouth
Basilosaurus head and mouth details
Not surprisingly, the head took especially much time to illustrate, as it includes the most detailed features. I followed the same considerations as in my earlier cranial reconstruction of Dorudon. The line and corner of the mouth were particularly tricky, and it took a long time and many changes until I was finally happy with it. Many reconstructions of archaeocetes shows them with a very deep corner of the mouth, what gives them a quite reptilian appearance. But this animals were mammals, and had after all still the same basic masticatory musculature as us. This includes the masseter muscles, which were quite well developed in archaeocetes. We can see quite well the original position and shape of this muscles from the zygomatic arches. Furthermore there were some other masticatory and facial muscles as well, which affected the shape and position of the corner of the mouth.
Lips
Basilosaurus with closed mouth. This works only with upper lips.
If you have read my earlier posts about archaeocetes, you will know that I am a strong advocate against the common shrink-wrapped reconstructions of archaeocetes with skull-faced heads. I think there are several reasons why we see this type reconstructions so often. People look at those nasty teeth in the jaws and simply assume that they must have been well visible in the living animal. The vague similarity of the skull proportions with those of crocodylians likely contributes to this as well. For exactly this reason we see nearly every year cases in which the rotten remains of odontocetes are mistaken for „prehistoric“ reptilian sea monsters, like this orca carcass from New Zealand.
Another important point is that artists usually like drawing teeth, and depicting an animal in a badass way is clearly preferred over a „lame“ look in many cases. I was actually surprised how awesome and badass those formidable Basilosaurus teeth still look in the final reconstruction, even with the upper teeth fully covered by the upper lips. It looks now much lesser than the usual crocodile-faced depiction with bare upper teeth. I think most people don’t realize that a lipless Archaeocete could never close its mouth. Even when its jaws would be fully closed the teeth would be still fully visible and the cavity of the mouth constantly in contact with the surrounding water. We see such a condition only in some few highly specialized modern cetaceans like the Ganges river dolphin or the pygmy and dwarf sperm whales. But nearly all other odontocetes have upper lips and cheeks that cover their upper teeth (except of course oddballs like narwhals, but the still have lips). Even odontocetes with large teeth like orcas and false killer whales have fully lip-covered upper teeth. We can be also quite sure that the most archaic and still amphibious whale ancestors had lips as well – like other artiodactyls and mammals usually do. It would be really weird if archaeocetes reduced and even lost their upper lips and their descendants re-evolved them later again. It is also surely important to keep in mind that the general jaw shape dentition was not even that different between the early amphibious forms and the fully marine archaeocetes like Basilosaurus. So lips surely did work around this arrangement and shape of teeth.
What’s just uncertain is the flexibility of those lips and when archaeocetes had still more flexible and perhaps partially retractable lips similar to seals and when already stiff lips like most modern odontocetes. Perhaps the later forms in-between had a somewhat similar condition as we see in beluga whales. I tried to place it somewhere in between.
This shows how the reconstruction was directly made over the original skeleton to get the proportions right.
Vibrissae
I also added some vibrissae on the top of the snout and upper lip, at top of the mandible and a few ones behind the nostrils. This are the areas where we still find vibrissae on modern baleen whales and in hippos as well. The whole topic of vibrissae in archaeocetes is quite complex, if you want to read more about it, take a look at this older article.
As a reminder you can see here the vibrissae of a North Atlantic right whale:
Vibrissae of a North Atlantic right whale (from Wikipedia Commons)
Nostrils
Take your fingers and touch your nose. Seriously, do it. Touch it on its back, at the base, the sides and the top. Move it. What do you feel?
You
will feel your Os nasale, the different cartilages on the sides and
on the tip, and you will also feel the tiny muscles which can use to
move the sides of your nose. We humans have somewhat weird looking
noses, but they have still the basic mammalian nose structure with a
lot of soft tissue over the skeletal opening of the nasal cavity. For
some reason comparably many reconstructions of archaeocetes simply
follow the outlines of the skull and only show some small openings
directly over the nasal cavity.
But
where are all the other parts of the nose? In odontocetes, we don’t
see much more than the blowhole as all the other parts of their
complex and asymmetrical nasal elements are hidden within the rounded
melons on the top of their cranium. The only exception is the sperm
whale which has a well visible lump of tissue around its blowhole
which is – unique in cetaceans – on the left side on the very top of
its giant spermaceti organ.
Skull of a blue whale, Centrum für Naturkunde, Hamburg
Baleen whales are a much better comparison with archaeocetes here. They lack the melons of odontocetes and have comparably little soft tissue on the top of their skulls. So the dorsal shape of the head is still quite close the the underlying skull – except for the fleshy hump which includes the two blowholes and the accompanying cartilages and muscles.
The fleshy nostrils of a breathing fin whale (Photo from Wikipedia Commons). Compare it with the blue whale skull above, there is really a whole lot of soft tissue over the opening of the nasal cavity.
Neck
The neck of archaeocetes was still longer and much more flexible than those of modern whales. But this still does not mean that the head and torso were as strongly separated as we see it in some reconstructions. I made the neck and throat similar to those of a seal, with a slight indication where the neck ends and the thorax begins. In odontocetes with quite flexible necks like belugas, snubfin dolphins or Amazon river dolphins we see also comparable shapes, which differs from those if other whales with stiffer necks.
Some reconstructions of archaeocetes have quite weird looking skinny necks which strongly separate the head from the body, what´s again not likely at all.
Flippers
The forelimbs of archaeocetes like Basilosaurus were still surprisingly underived compared with the flippers of modern whales and even the forelimbs of seals. They are comparably long and show neither really strong attachments for muscles nor elongations of the fingers. The reason is possibly the fact that archaeocetes had not that much use fore their forelimbs. The basal amphibious forms used their strong hind legs and tail to swim and maneuver underwater, similar to many modern amphibious mammals like otters, beavers, nutrias or even desmans and water opossums. In those animals we find often well developed webbing of the hindfeet, but much lesser on the fore feet.
Forelimbs of a Carribean manati, photo from Wikipedia
We see the probably most derived adaptations to a marine habitat of a modern „walking“ mammal in the sea otter, which spends nearly its complete life in the water. This surprisingly large durophagous mustelid has only very short arms, whose main function is to find and collect invertebrate prey, and to crush open hard shells and sea urchins with stones. So I decided to reconstruct the forelimbs not as propper flippers but more like the comparably stumpy and leg-like arms of seacows. I also added a more speculative feature, some vestigial nails. I think so far there was no research to examine the proximal phalanges of archaeocetes to see if there is any indication for such nails. Their superficial structure of the bone could probly solve this mystery, to see if there was still any keratinous growth or not. Seal still have claws, which can be pretty big. Even the highly modified fluke-shaped hind limbs of phocids have short nails. In seacows, which are even fully aquatic we see small hoof-like nails in their forelimbs as well. So even if we don’t have direct evidence for vestigial nails in archaeocetes, we have also so far no direct evidence against it, and the idea really does not seem too far-stretched. So I gave my Basilosaurus seacow-styled protoflippers.
Tail and fluke
Basilosaurus tail and fluke reconstrucion. Adding scars to indicate some things which happend in the life of an individual is always fun.
The original version of this Basilosaurus had still a deeper tail and a more conventional shape of its fluke. But I decided to reduce the tail somewhat and make it a bit slimmer, as the dorsal spines of the caudal vertebrae were still comparably short in Basilosaurus. As it also still lacked the caudal peduncle of modern cetaceans I tried to give the caudal fin a somewhat lesser derived shape. The flukes of archaeocetes are really tricky as we can’t reconstruct their shape from the underlying tail vertebrae as in ichthyosaurs and certain mosasaurs or metriorhynchids. Of course we can take a close look at the last caudal vertebrae und see if there’s something going on with them, for example if they are wider and more robust than the vertebrae before them. But this still doesn’t give us really much more information about the original shape of the tail or fluke. Archaeocetes started quite early to use their tails as primary propulsion and their forelegs were likely mainly for maneuverability whereas their hind legs were already fully useless for swimming.
This
indicates that a caudal fin was already present in the very early
fully marine archaeocetes. But how did it look like? It surely
started as a similarly dorso-ventraly flattened tail as those we see
in otters and there was also quite likely a stage with a primitive
spoon-shaped tail like those of manatees. But the problem is that
even with a totally complete line of evolutionary stages between the
tail vertebrae of the most basal archaeocetes up to those of modern
cetaceans with high dorsal spines, laterally compressed tails and a
caudal peduncle for the fluke, we would still don’t know how the
caudal fin really looked like.
To get a better understanding of this issue we can take a look at seacows. The manatees which mainly populate freshwater habitats in slow moving rivers or coastal areas (but don’t forget, they also crossed the Atlantic ocean anytime) have only a fleshy oval caudal fin, but it seems to work obviously well enough for them. Dugongs which are living in marine habitats are much more affected by turbulences, waves and similar things and have already a nearly whale-like fluke. They spread over large areas of the globe, over Africa, Asia and Australasia what indicates that they are quite seaworthy. In contrast to manatees they are also much more affected by predation, for example by tiger sharks. In the past they were quite likely also hunted by predatory cetaceans and in some areas possibly also by large marine crocodylians.
This are possibly the reasons why dugongs appear much more whale-like than manatees, as there was a much harder selection for strong swimming abilities than for manatees. Their caudal vertebrae also show elongated dorsal spines and some flattening of the most proximal vertebrae, but we would probably not expect such a well developed fluke from the bones alone. In direct comparison with those of whales it’s however still somewhat lesser derived. The tail is still comparably wide and the fluke itself comparably thick and fleshy.
Caudal vertebra of a dugong, Staatliches Museum für Naturkunde, Stuttgart
This could give us some insights into the evolutionary history of whale flukes. I could imagine that there also was a strong selective pressure on the shape of archaeocetes as already little differences of its structure could have had comparably strong hydrodynamic effects. And we also shouldn’t forget that archaeocetes were unlike manatees or dugongs no herbivores but predators which hunted fast and agile prey. That’s why I tend to assume that highly derived oceanic predators like Basilosaurus already had comparably well developed flukes. So I reconstructed the caudal fin as a real fluke and not like manatee paddle, but still different from those of modern cetaceans. It’s still thicker, somewhat longer and more rounded. My very first version was still closer to the flukes of extant cetaceans but I wanted to keep the differences in vertebra shape more in mind and make this also somewhat more obvious in the external shape.
I also gave my Basilosaurus no dorsal fin. One reason for this was because I wanted to make a bit similar to right whale dolphins, little known odontocetes with a very thin and long body, which fully lack a dorsal fin. Another reason was to depict the Basilosaurus as different and more primitive compared with modern whales. However, we have to keep in mind that baleen whales and odontocetes diverged already very early in cetacean evolution, and both do have dorsal fins. So this means that those most basal ancestral baleen whales had either already a dorsal fin (perhaps a very primitive one at least) or that dorsal fins evolved later parallely in both lineages. That´s some food for thought.
Coloration
The coloration of prehistoric animals contains for obvious reasons an especially big amount of speculation, unless we have cave paintings or exceptionally well preserved fossils which can give us some direct evidence for the patterns and colors. In the case of Basilosaurus we have at least some extant relatives, but they can give us still only a vague hint about the colors and patterns of prehistoric whales. I wanted to avoid just another Basilosaurus with the colorations of the Walking with Prehistoric Beasts Basilosaurus. It really doesn’t look that bad and it’s not implausible but I just wanted something different. I’ve discussed the coloration of large marine animals for the paint job of the Meyerasaurus by my friends of kamyk.pl for the museum at Lodz at length here. For the Basilosaurus I also made a first version with a similar color scheme that was somewhat reminiscent to those of a leopard seal. Then I tried a more homogeneous and darker main color which was inspired by false killer whales and long-fin pilot whales, which have also quite elongated bodies. I also made no classical counter-shading with a generally lighter underside. This is more a trend than a rule and a large number of marine tetrapods, especially seals but also a lot of whales, show no real counter-shading at all. Instead I made the area of the throat visually distinctive with a white patch, something we also find in many different modern whales. Because of the incredibly elongated body of Basilosaurus I made also a small white patch in the area of the small hindlimbs. I could imagine that such an eyecatcher of the genital area could make sense in an animal which had possibly really some obstacles to find the right position during mating. We find also similar white eyecatchers (or, as I call them, cetacean arseantlers) in a lot of very different cetaceans, from right whales to sperm whales and various other odontocetes. This brings us to the next point.
Sexual dimorphism
One of the great things about digital painting is that you can recombine your work again and again. So I made just another Basilosaurus from the first one, what gave me also some more good chances to depict some other aspects. At first I scaled it down to show the sexual size dimorphism. Male Basilosaurs were about 20% longer than females, based on studies of their bones. The second Basilosaurus was also a good opportunity to depict it with a closed mouth. Its mouth is now fully shut and the lips make the teeth no more visible. Remember this couldn’t work at all without upper lips, the teeth would be exposed all the time.
Pair of male and female Basilosaurus
To make the two somewhat more individual I changed the position of the forelimbs of the male to indicate a certain approaching position towards the female. The male has also a slightly lighter coloration, something we also see in older males of many cetaceans like sperm whales or bottlenose whales. He received various scars as well to indicate some intraspecific fights with other males. This is also a quite common thing in various odontocetes from sperm whales to many beaked whales and a lot of dolphins. Last but not least he got a genital organ as well. In fact this was the very first time that I have ever depicted something like this. But prehistoric animals were not always just fighting, hunting and eating and even a gigantic apex predator like Basilosaurus had a social life and sexual interaction. One could even say that next to everything an animal does, no matter if it’s hunting, eating or fighting, has the main purpose to ensure that it can successfully reproduce during its life. So here we go, Basilosaurs having fun.
Gosh! – Archaeocete sex!
Background fauna
You shouldn’t underestimate the background fauna. Sometimes it can be somewhat annoying and time-consuming to paint it, but if you have already spent a lot of time on your main subject you can also give it some nice substitutes. You should see the background fauna as a chance and not as a time waster. It gives you a chance to depict some interesting and possibly hardly ever shown species with comparably little effort (unlike a full life-depiction of those animals being in the focus themselves). For this reason you should also avoid totally random stereotypes but really take some time to make a correct depiction. Fish are quite typical elements for background or „prey“ fauna but unless you make a swarm of really tiny and individually unrecognizable fish you should really try to make a realistic depiction. If you are not really familiar with the animal you want to depict, you will very, very easily make errors. I write this because that’s exactly what happened when I started with some unspecified random fish. So I was searching for something better and decided to make a swarm of mackerels. They have a very good fossil record with great specimens from the Eocene which were already very similar to the modern forms. They would also fit within a scenery in the open water and their familiar appearance gives also some more scale for the size of the Basilosaurus. Furthermore I could depict a visually more interesting fish than just one with silver-white colors.
Mackerels
There
are also some recycled nautiluses in the background to make the fauna
a bit more diverse and to add something that appears somewhat
ancient and primordial.
I spent also a considerable amount of time on a Dorudon which was originally only planed as a simple background illustration but at the end it became a similarly detailed reconstruction as the Basilosaurus itself. At first I wanted to depict Basilosaurus hunting a juvenile Dorudon, but then I wanted to depict it next to an adult specimen. The skull length of Basilosaurus and Dorudon is surprisingly similar, despite the very different total lengths. Those of Dorudon has however more elongated jaws and is lesser massive than those of Basilosaurus. But this was still a pretty big animal, about as large as a false killer whale, and with really large jaws.
Basilosaurus hunting Dorudon
The Dorudon in detail:
Dorudon reconstruction
What I really want to emphasize here is that I don´t think this particular reconstruciton is the one and only possible. That´s why I wrote where things were more speculative but also why I made it the way I did. Some traits like the upper lips and the vibrissae (perhaps lesser pronounced, but perhaps even more) were nearly certainly present in archaeocetes, whereas other traits like the shape of flippers and fluke or the presence of a dorsal fin are much more of an issue.
References:
Gingerich, P. D., B. H. Smith, and E. L. Simons. 1990. Hind limbs of Eocene Basilosaurus isis: evidence of feet in whales. Science, 249: 154-157.
Zalmout, Iyad S. 2000. Priabonian Basilosaurus isis (Cetacea) from the Wadi Esh-Shallala formation: First Marine Mammal from the Eocene of Jordan. Journal of Vertebrate Paleontology 20(1):201–204.
It is well known that there are a lot of wonderfully preserved ichthyosaur fossils with skin impressions, especially from the famous Posidonia shale of Holzmaden. But it is much lesser known that there are also some soft tissue impressions of other marine reptiles as well, for example of some plesiosaur specimens. Admittedly, so far none of them is as well-preserved as the best ichthyosaurs fossils. But today I don´t want to show you ichthyosaurs or plesiosaurs but a truely spectacular fossil from the Posidonia shale of Frittlingen. It is a remarkably well preserved leg of a Steneosaurus bollensis, a medium sized marine crocodile which is next to the ichthyosaurs among the most common marine reptiles of the Posidonia shale.
This particular fossil is neither very big nor complete, it´s just a single hindleg, nothing you would normally show in an enormous paleontology museum like the Museum am Löwentor at Stuttgart, which has so many bigger and more complete Steneosaurs in its collection. But it´s still really special, as this leg has not only its bones preserved, but also parts of its skin. I had to make a composite of two different photos to show you the whole leg at a better foto quality, as it was hardly possible to take a photo of the whole leg without getting reflections.
Steneosaurus bollensis, Posidonia Shale Frittlingen, Museum am Löwentor Stuttgart
In some respects this fossil is even more interesting than those of ichthyosaurs, as we can still see really fine surface structures and patterns of the skin, even with individual scales. You can see a more detailed photo of the upper part here:
And of the lower part:
Of course you want to see some more detailed photos of the scales, which are very small and rectangular in shape. This is likely the inside or underside of the leg, where we find also smaller scutes in modern crocodiles.
Compared with the skin of modern crocodylians, they are still very fine and numerous. Here is a photo which I took at Berlin Zoo, which shows the differences in size and arrangment of the scutes between the outside and inside of the leg:
Hindleg of a spectacled caiman (Caiman crocodilus), Berlin Zoo
Here a direct comparison:
Spectacled caiman (Caiman crocodilus) leg compared with Steneosaurus leg
If you take a close look, you can still even see where the scales had wrinkles in-between:
Skin folds between the scales
This is really quite interesting as it seems to indicate a trend toward a fine, more flexible and smoother skin in marine crocodylians, which had clearly a very different biology than all crocodylians we see today. I write „seems to indicate“, because it could be theoretically also possible that the ancestors of Steneosaurus already had finer scutes than extant crocodylians. A simplifaction and smoothing of the skin would be however fully plausible within a lineage which evolved into mainly aquatic and quite likely much more agile marine predators. I find it quite interesting that among all modern crocodylians we also find the smoothest and least gnarly skin in the most aquatic form, the bizarre gharial (Gavialis gangeticus). Even its dorsal osteoderms are much lesser pronounced than in crocodiles or alligators.
Gharial at Berlin Zoo
Even their limbs look much more reduced and paddle-like than those of other modern crocodylians.
Gharial forelimbs. Not the smooth surface and the stumpy fingers.
We also find a strong reduction of scales in highly aquatic turtles like pig-nosed turtles, softshell turtles and the leatherback turtle, and we also know that mosasaurs had only tiny scales, and ichthyosaurs even no more scales at all.
It would be really interesting to know how the skin looked in the most derived marine crocodylians like the metriorhynchidae, that included large macropredatory forms like Dakosaurus maximus. Fossils like this Steneosaurus leg are of course also extremely worthy for paleoartists, as they show us information about anatomical details which we could never know for sure from the bones alone. Let´s hope that more of such superbly preserved fossils will be found in the future.
Today I want to feature some paleoart by Dominic Grabowski, which depicts the archaic stem-baleen whale Mammalodon attacking one of those awesome but little-known giant post-creataceous nautilids. I feel really honoured that this was directly inspired by my recent blogpost about post-cretaceous nautilid diversity which I wrote some time ago. This means that my work here can really influence other people´s paleoart and encourage them to deal with certain subjets, and that the countless hours I spent to write my blogposts are appreciated. Thanks Dominic!
Mammalodon attacking nautilid by Dominic Grabowski
If you are interested in unusual freshwater fish, you have possibly heard about the taimen (Hucho taimen), a very large predatory salmonid which lives in remote parts of Mongolia, Russia and some other areas of North Asia. This species was already covered in an episode of River Monsters with Jeremy Wades and also in the National Geographic Monster Fish series with Zeb Hogan. The habitat in exotic and remote areas outside the western world clearly increased the nimbus of the taimen as an elusive monsterfish. It is much lesser known that the taimen has a close and nearly equally impressive relative which lives even right in the centre of Europe.
Adult huchen (Hucho hucho) at the river Drina between Bosnia and Serbia. Photo from Wikipedia
The name of this fish is Huchen (Hucho hucho), sometimes also still called Donaulachs (Danube salmon) in German. Despite this old name, the huchen is no real salmon. The genus Hucho represents together with the related Brachymystax genus a unique clade within the Salmoninae subfamily of Salmo, Oncorhynchus, Salvelinus, Parahucho and Salvethymus that includes the very diverse and numerous trouts, salmons and chars. For such a big predatory coldwater fish it is surprisingly colourful, with copper-red flanks at the back-half of its body. It has also an unusually large adipose fin.
Juvenile Huchen of about 40 cm, Wilhelma Zoo, Stuttgart. It lives with several other huchen and some brown trouts in a big paludarium.
The huchen lives in fast-streaming water, and has a very elongated and spindle- shaped body. In cross-section, most of its body is nearly round, unlike those of trouts, salmons or chars, which have an oval cross-section. The head of the huchen is comparably flat with a very wide mouth, and its jaws and tongue are equipped with well developed teeth. In contrast to many trouts, salmons and chars, male huchen never grow a hooked mandible during the mating season.
Mouth of a taxidermy huchen, Haus der Natur, Salzburg.
It is a voracious predator which feeds on other river fishes like the common nase. It is also well known to attack tetrapods on occasion. Small rodents and small birds like kingfishers or dippers were already found in the stomaches of huchen. For this reason, baits which imitate swimming rodents are quite popular huchen lures. In earlier times lampreys were also an important component of their diet, but those primitive vertebrates have become quite rare today. There is even a traditional bait called the Huchenzopf which imitates a group of small lampreys. Old models were usually made from a lead weight and pieces of leather or rubber tubes, today the are usually made with soft rubber.
The original range of the huchen is restricted to the fast-streaming and clean tributaries within the basin of the river Danube, as well as a few lakes like Lake Plav at Montenegro. There were also unsuccesful attempts to introduce it to rivers in France, the Netherlands and Great Britain. Surprisingly such artificial stocking was however successful in some rivers at Spain and Morocco, but those populations are not sustainable. According to one single source I could find, specimens from the Bulgarian huchen populations occasionally swim into the coastal waters of the Black Sea. But I could sadly find no further information about this. It would be extremely interesting to know if this is really true, and if huchen show also similarly strong morphological changes in colour and body shapes as sea-dwelling brown trouts, rainbow trouts or chars.
The huchen also lives and even breeds in the river Isar, even right within the city of Munich. That´s really an unusually non-exotic home territory for such an enormous and generally little known near-megafish. Just keep this in mind should you ever go visiting Munich. You´re walking there alongside a river inhabited by the second largest non-anadromous salmonid of the world.
River Isar at Munich
The huchen is usually the largest predatory fish of the fast-streaming and comparably cold waters where it lives. It reaches often lengths of over a meter, and record specimens can reach at least lengths of 1,30 –1,44 m and weights of around 30-40 kg. You can see some photos of exceptionally big specimens here. Of course there are as usual stories of bigger ones, but as big-fish-stories are notoriously untrue and the largest claimed sizes for fishes are quite often unconfirmed anecdotes and hardly trustworthy, I prefer to stay on those lengths for which we can find confirmed data. But even then, the huchen is a huge fish. Among the non-anadromous fish of middle Europe, only the wels catfish grows considerably bigger. It also exceeds the largest ecotypes of lake dwelling brown trouts (Salmo trutta) and even the official angling record specimens of the northern pike (Esox lucius). Only the very largest Atlantic salmons (Salmo salar) reach similar record lengths and even higher weights, like the historical monster specimen covered some time ago. But this salmonid is anadromous and gains its size in the sea. For a pure freshwater fish which lives in comparably shallow and cold streams, the huchen is a true giant.
Here is a Huchen of 130 cm lenght and a weight of 25 kg, 125 g which had a 79 cm girth. Caught in 1961 in the river Loisach near Eurasburg. Photo taken at Deutsches Jagd-und Fischereimueum (museum of hunting and fishery) Munich.
Sadly – and not really surprisingly- the huchen isn’t doing well. As many other fishes which need clean and oxygen- rich streaming water, it suffered considerably from pollution and changes of the original shape of the streams. One of the most serious problems is the loss of most of its spawning grounds. The huchen needs clean gravel in fast streaming and clean water for its spawn. But such areas have significantly decreased in number. Another problem is the decrease of prey fish for the young huchen, like the common nase (Chondrostoma nasus), a cyprinid which populates the same streams as the huchen. Its fry is among the most important prey items for small huchen.
A super-bulky and testosterone-bloated male brown trout which was rubbing itself for a considerable time on the back of a subadult Huchen. Photo from Wilhelma Zoo, Stuttgart
Today the huchen can only survive in many streams due to artificial stocking. It is a good thing that this works at all, and that a few adult specimens can produce vast numbers of eggs which can be used to breed thousands of young fishes. But the other side of the medal is a massive reduction of genetic diversity in those fish. That is one reason why it is so important to make sure that specimens of endangered species should be able to survive and reproduce in the wild.
In terms of zoological diversity, the Baltic Sea is probably the most boring sea of the world. It has the lowest salinity of all seas and it is only connected with one other sea, the North Sea. As a result of this it has only a very low number of native species, and those are also not particularly spectacular compared with those of the North Sea or the northern Atlantic.
What’s especially noteworthy is the nearly total absence of any bigger animals there. The largest marine mammals which are really native there are grey seals, harbor seals and harbor porpoises. The grey seal and thr harbor seal were already nearly fully extinct in the late 20th century and today there are only a few hundred specimens left. The harbor porpoise is also extremely rare and among the smallest cetaceans of the world. So even the mammalian marine megafauna is not exactly impressive to say it at least. As a result of the low salinity no sharks are native in the waters of the Baltic Sea. The biggest commonly occuring fish you can hope to see are only Atlantic salmons, sea trouts and cods. All of them rarely grow bigger than a meter as even the cods of the Baltic Sea stay much smaller than their cousins in the Atlantic Ocean and North Sea. Of course there are occasionally cases of vagrant specimens of bigger fish which usually don’t occur there, or even out-of-place cetaceans like rorquals or sperm whales. But this are rare exceptions and they could not even survive for a long time in the Baltic Sea, as they could not find suitable prey in this shallow sea. As you see, aquatic megafauna is next to absent there.
This makes it even more surprising that there actually were gigantic fishes in the Baltic Sea until just a few decades ago. Giant blue fin tunas were common denizens of the Baltic Sea. This predators are among the largest teleost fishes of the world, with record specimens of more than 650 kg and lengths of more than 3,5 m. The ones caught in the Baltic Sea were not that gigantic, but still dwarfed every other fish around, since the European sturgeon which also once populated this waters has become practically extinct in the early 20th century. Some of the tunas living off Denmark grew to massive sizes, like this one which was caught around 1880 whose skeleton is now on display at the zoological museum of Copenhagen.
Skeleton of Atlantic bluefin tuna from around 1880, Zoological Museum Copenhagen
Sometimes tunas of more than 300 kg were caught, and the record was even a massive specimen of 372 kg.
Here is skull of a big tuna caught in 1962 at Howachter Buch, Germany, now on exhibit at the zoological museum of Kiel.
Bluefin tuna skull, Zoological Museum Kiel
I added a pocket rule of 50 cm to give you a better idea of the size. Here is another photo which shows the skull with the reconstructed outline of the whole fish.
Bluefin tuna skull, Zoological Museum Kiel
Bluefin tunas were regularly caught by fishermen in Danish and even German waters. They were so frequently caught that a factory for canned tuna was built in Skagen in 1929. Sport fishermen came even from Great Britain to Denmark for big game fishing. But the whole business ended in the 1960ies when the tunas disappeared from the Baltic sea. The reason was overfishing, not only in Denmark but also in other parts of the world like in the spawning grounds in the Mediterranean Sea and the Atlantic ocean where the bluefin tuna has its nursery ground.
Today there is not much that reminds on this giants anymore. At Odden Havn, a small fishing port at the north coast of Sjaelland, you can still find old photos from the time of commercial tuna fishing, which seem nearly surreal today. Here is a historic photo of fishermen with tunas from Odden Havn, Denmark.
This port was one of the former centers of tuna angling and fishing. Even today there is still a tavern name tuna. Many years ago, there was still a large taxidermy head and caudal fin of a trophy tuna at the „Tunen“-restaurant of Odden Havn. I think it is now on exhibit at the tuna fishing museum of Odden Havn, which opened just a few years ago. Sadly it was every time closed when I was there, even at days when it should have been open, so I don’t have any photos from the interior.
The tuna museum at Odden Havn, Sjaeland.
The old trophies, skeletal remains and black and white photos seems to be the last remnants of a bygone era. But in recent years, the first blue fin tunas came back to the Baltic Sea. In September 2017 a swarm of tunas wad sighted off and subsequently a project was started to tag some of them. One of them was caught with hook and line, a huge specimen of 2,51 m length and an estimated weight of 285 kg. This was the largest fish which had been caught in this waters since half a century. After it was tagged, it was released again. Of course the tunas are now strictly prohibited there and catching them is only allowed for scientific research.
The „Tunen“ restaurant at Odden Havn.
It will likely take many decades until tunas are again regularly found in the Baltic Sea, but their eventual reappearance seems to be a good start at least. But it is also a reminder that tunas are not just big tasty fish but endangered marine wildlife which are already exterminated in much if their original range. They are marine megafauna like cetaceans, seals or sharks, but they still don’t get the same sympathies. Even people who are outraged about the slaughter of whales for „scientific research“ or the killing of sharks for their tasteless fins will often still happily and regularly buy and consume tuna. I strictly reduced my tuna consumption already many years ago and it would really help if more people would appreciate those fish not just for their culinary qualities but see them instead as endangered wildlife and giant marine predators which are close to the top of the food chain.
To conclude, I really have to apologize for my seemingly harsh words towards the Baltic Sea at the beginning of the blog. I absolutely love the Baltic Sea, it is by far the sea with which I am most familiar with, and I it has its very own loveliness and beauty.
This blog is devoted to the wonders and marvels of the animal kingdom and natural history. Many of the photos in my articles were taken in zoological or paleontological museums. But you can also often find a lot of really interesting zoology-related artifacts in other museums as well. I have a longstanding interest in archeology, history and ethnology, and for that reason I visited countless museums about those fields over the years.
I have seen there innumerable and sometimes really surprising exhibits which can be also of great value for people interested in zoology. Not everyone can spend several hours analyzing the construction of ancient arms and armor or the carving techniques of old wooden spoons as I sometimes do. But even if you are not really an avid fan of archeological, historical or ethnological museums but interested in animals (I suppose you are, otherwise you would not read my blog), I can encourage you to take a closer look at those locations. I think I can not emphasize enough the potential of such collections, and sometimes you can even find amazing and fully unexpected zoological treasures that you will rarely find even in natural history museums.
The vestigal narwhal tusk featured recently is such a case. The Inuit tools made from the teeth of Greenland sharks were the very relics of Somniosus I have seen anywhere. Another example would be the mandible of a pygmy sperm whale in the Oceania exhibition of the ethnological museum at Berlin (I already wrote about it several years ago here).
Pygmy sperm whale (Kogia breviceps) mandible, Ethnological Museum Berlin
The mandible was sadly labeled as the jaw of a dolphin. I tried to contact the museum to inform them about the true identity and rarity of this mandible, which belongs to a species which is really rare even in the collections and archives of museums with many cetacean specimens. Sadly I received no answer.
This leads to another thing. This is really not meant to discredit the scientists, curators or the staff of those museums, but over the years I have seen a whole lot of erroneously labeled exhibits in archeological, historical and ethnological exhibitions. For this reason it can be really beneficial if people with a more profound knowledge of zoology take a closer look at them. Of course no one likes a smartass (ans yes, I am fully aware of my strong tendency to be one), but museums have an educational purpose and labels should be corrected if they are erroneous. At the end, that’s also beneficial for those museum and their visitors, in particular if a „boring“ object becomes suddenly much more interesting.
This is even more important if erroneous identifications lead to erroneous interpretations, what’s especially problematic in the archeological context. I hope to feature such a case in the near future.
But enough with the criticism, in the vast majority of cases the identifications of animals in such museums are correct, an you can learn a lot from them.
Real physical remains of animals are of course especially interesting, the more so if they belong to unusual or now extinct species. I have seen many subfossil aurochs skulls and horn cores, and even some full skeletons, and a great number of them in archeological museums. This included even a particularly spectacular (perhaps even the most spectacular) specimen which I will spare for a future blog post. So if you are a fan of Bos primigenius, you can likely expand your specimen count in archeological museums to a substantial degree.
Aurochs (Bos primigenius) horn cores (on the top) in the archeological museum Strasbourg. Did you notice the warthog model?
But even the skeletal relics of lesser uncommon animals are always worth to take a closer look at them, no matter if they are wild or domestic animals. Sometimes you will also see bones of animals which are now extinct in the area where their relics were found, like Central European mooses. Some animals which greatly suffered from trophy hunting over many centuries have also often decreased in size. For that reason you can often find especially massive deer antlers or huge wild boar tusks which come from archeological excavations or which are from historical hunting collections.
A huge neolithic wild boar skull with enormous tusks from the National Museum of Denmark, Copenhagen
Ethnological collections are sometimes real treasure chests, as they often include really exotic animal remains. Anatomy, behavior and ecology of animals are all fascinating things, but I think the interaction of animals with humans are always important to look at as well. The cultural or economic role or value of animals, the way in which they were hunted or caught or what was made from them. This are all really interesting topics of themselves, but they often get not that much attention in zoological museums.
Necklace made from hornbill beaks, likely from New Guinea. How often can you take such a good look at the inside of their mandibles? Photo taken at the Ethnological Museum Munich
You are sometimes really surprised by the weird hunting methods invented for certain species or the curios use of certain animal parts. Sometimes such processed body parts can give you even insights into anatomy that you will usually not see at a taxidermy specimen or a mounted skeleton.
A very nice collection of Crocodylus porosus skulls and boar skulls. Ethnological Museum Munich
In exhibitions about Oceania you will often find especially many artifacts made from animals.
Necklace made from sperm whale teeth. This gives you a pretty good idea about the variation in shape of sperm whale teeth. The big blunt tooth is quite interesting, as it shows severe abrasion. Photo taken at the Ethnological Museum Berlin
Native American artifacts can be also very interesting, for example if they give you a chance to take a close look at bison skulls.
Bison skull with painted ornaments, Ethnological museum Berlin
But not just physical remains can be valuable objects of interest, but artistic animal depictions as well. In some cases they can tell you things about now extinct animals that you can’t see if you just look at their bare bones. Colours, patterns, even certain typical kinds of behavior and interaction with other animals and humans can be found in ancient art or historical depictions.
Mammoth ivory figurine of a cave lion from Vogelherd Cave at Schloss Hohentübingen. Could the pattern carved on the body indicate that some cave lions had more or less pronounced spots?
One especially worthy field to discover in archeological museums are old forms of domestic animals. You can for example look which unusual traits typical for domestication were already present. You can take a look at ear shape, length or structure of the fur, the pattern and colour of the hides or other traits you can not judge from skeletal remains alone. Many old breeds of domestic animals became fully extinct, others changed so much that they bear no more much similarities with their ancestral forms.
A greek bronze helmet from 300-400 BC. The engraving of the bull is just wonderful and incredibly naturalistic. It reminds a lot on a Spanish fighting bull.
A small roman bronze figurine of a domestic bull with strongly developed dewlap.
Roman bronze bull, Landesmuseum Stuttgart
Terracota model of a South American dog breed. This small, short-legged dogs had strongly wrinkled skin and were bred for their meat.
Ethnological museum Berlin
Domestic dogs were already quite diverse in antiquity, and very different breeds already existed millenia ago.
A greyhound-like breed of dog on a Greek amphore, ca 440 BC. Photo taken at Landesmuseum Stuttgart
You can for example see typical traits of domestication in some of those old breeds, like floppy ears of a curled tail.
Dog with floppy ears and curled tail on Greek amphore, 480-470 BC, Antikensamlung Berlin
A very nice life-sized sculpture of a hunting dog from ancient Greek. This is in fact a cast from gypsum and not the original.
Greek hunting dog, Schloss Hohentübingen
Of course you always have to keep in mind that there can be a lot of artistic freedom in such old depictions. Sometimes more, sometimes lesser. But to get a better idea and understanding of that, it is important to study the various art styles of different cultures. In some cases you will be really surprised by the level of accuracy of those ancient artists. Some of the ancient Egyptian paintings and reliefs are so naturalistic that it is easy to identify even a lot of animals to species level.
On occasion you will even see quite unexpected scenes of human-animal interaction, like apparent attempts to tame wild animals. It is also sometimes really surprising to see the degree of human translocation of animals which happened already hundreds or thousands of years ago.
How to transport an elephant on a ship, Roman mosaik from Veji, 300-400 AD, Landesmuseum Karlsruhe
The depicted elephant was most likely the now extinct small North African subspecies Loxodonta africana pharaohensis.
You can also sometimes see unusual curiosities from historical collections, like anomalous antlers, horns or tusks, or bezoars which were thought to have healing powers, similar to the inevitable narwhal tusks. In other cases there are very early examples of exotic animals or animal parts which found their ways into cabinets of curiosities or early zoological collections. You will find such collections much more often in historical than zoological museums, and they can give you interesting insights into the history of modern zoology.
Bird of paradise in historic cabinet of curiosities, Landesmuseum Karlsruhe
Here you can also see a wonderful mug made from narwhal tusk. You can see that was from the base of the tusk, as the surface is unabraded and highly textured.
Narwhal tusk mug, Museum für Kunst und Gewerbe, Hamburg
An original copy of Conrad Gessner´s (1516-1565) Historia animalium, one of the very first zoological books of the world. The same exhibition also had an original print of Dürer´s Rhinoceros.
Historia animalium by Conrad Gessner, photo taken at a special exhibition at the Museum für Kunst und Gewerbe, Hamburg
I hope I could show you with this blogpost why it can be always worthy to think outside the box, and to visit as many museums as you can, even if they are not always restricted to natural history alone.
Diese Website benutzt Cookies zur Identifikation von Benutzern und zur statistischen Auswertung von Inhaltsaufrufen. Mit der Nutzung erklären Sie sich damit einverstanden.












 Last year I was involved into a fantastic project, the production of the wildlife documentary „Into the Forest: Reptiles & Amphibians“ by my good friend Bryan Maltais.He made already several other shorter documentaries, including the award-winning production „
Last year I was involved into a fantastic project, the production of the wildlife documentary „Into the Forest: Reptiles & Amphibians“ by my good friend Bryan Maltais.He made already several other shorter documentaries, including the award-winning production „

































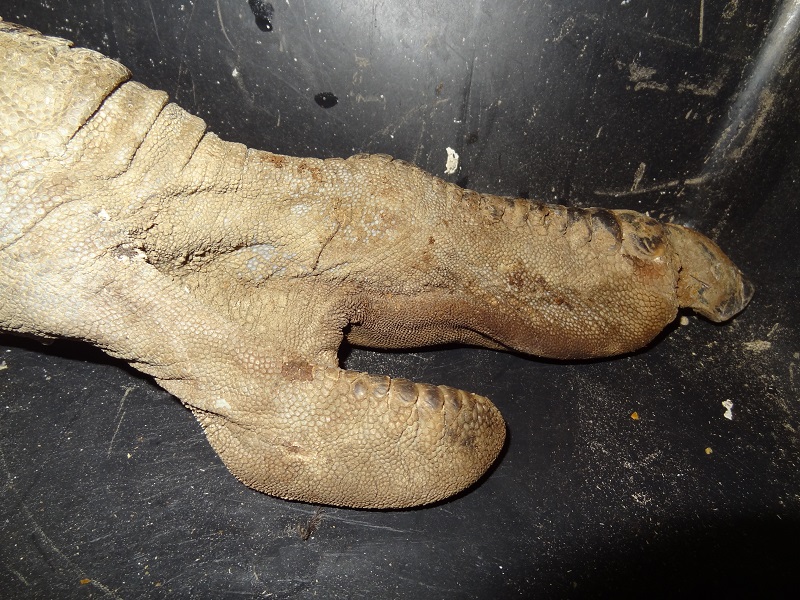
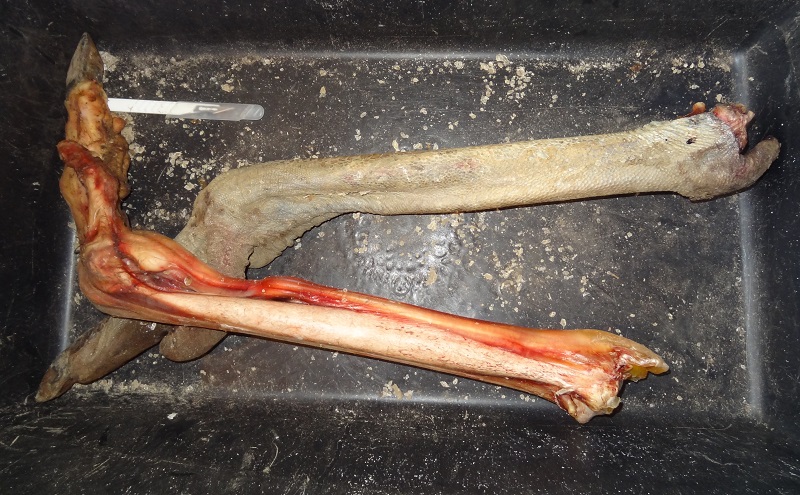


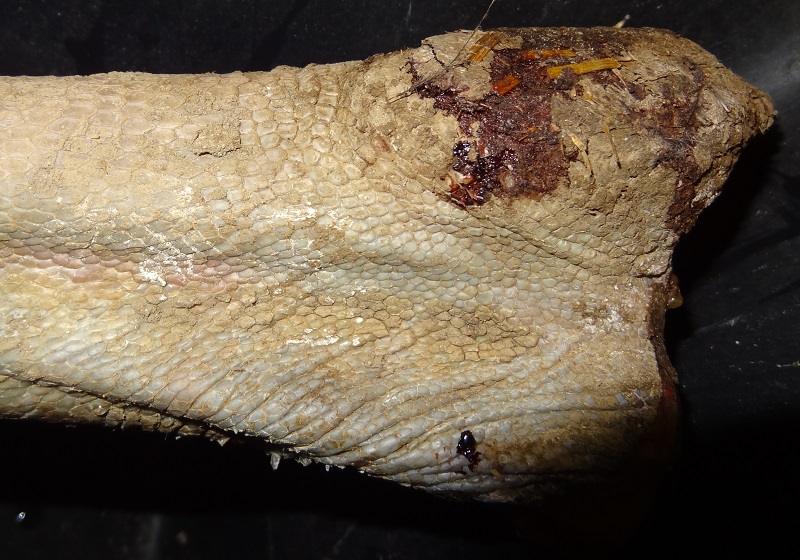




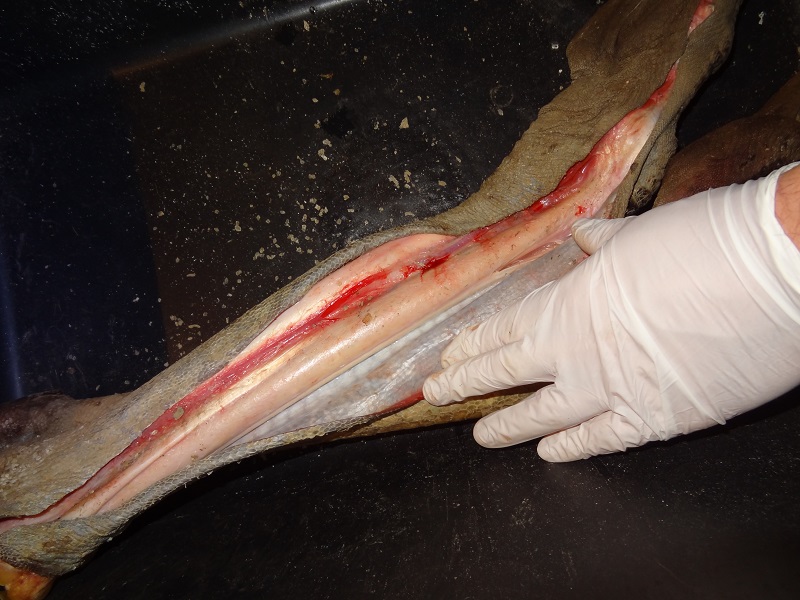








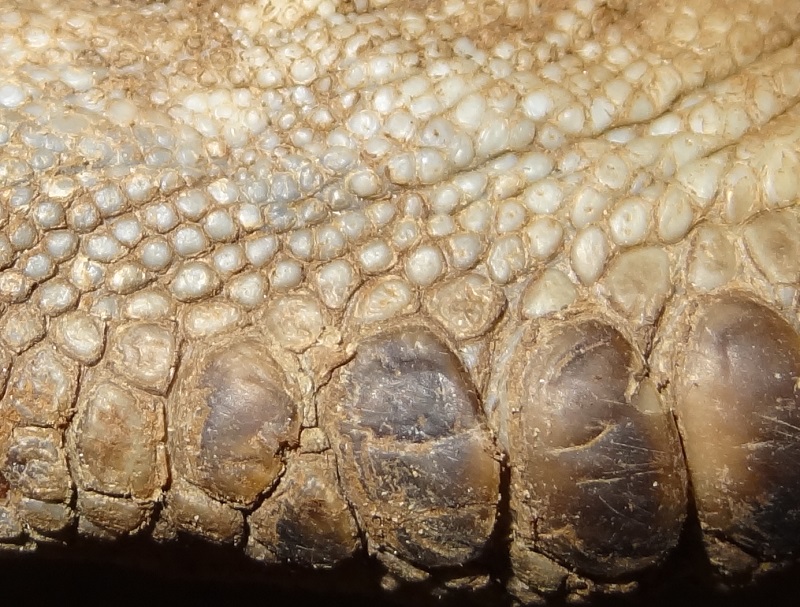


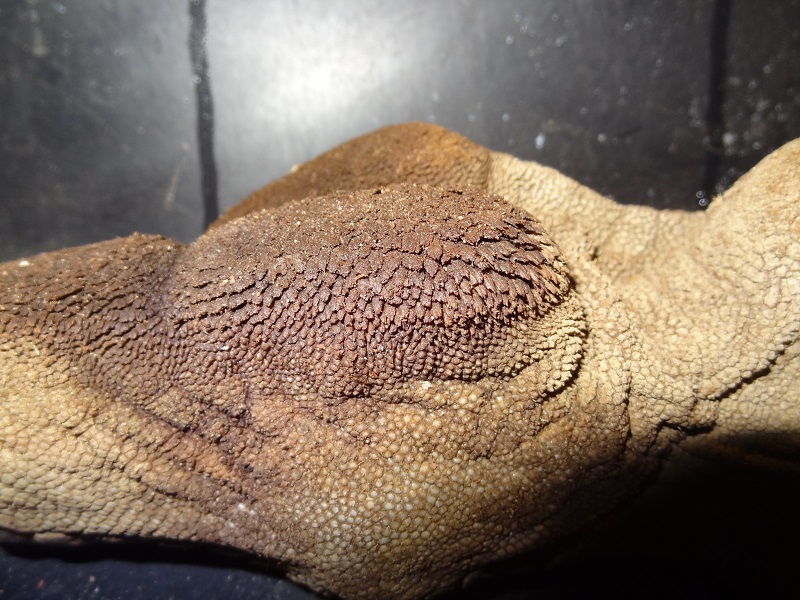




















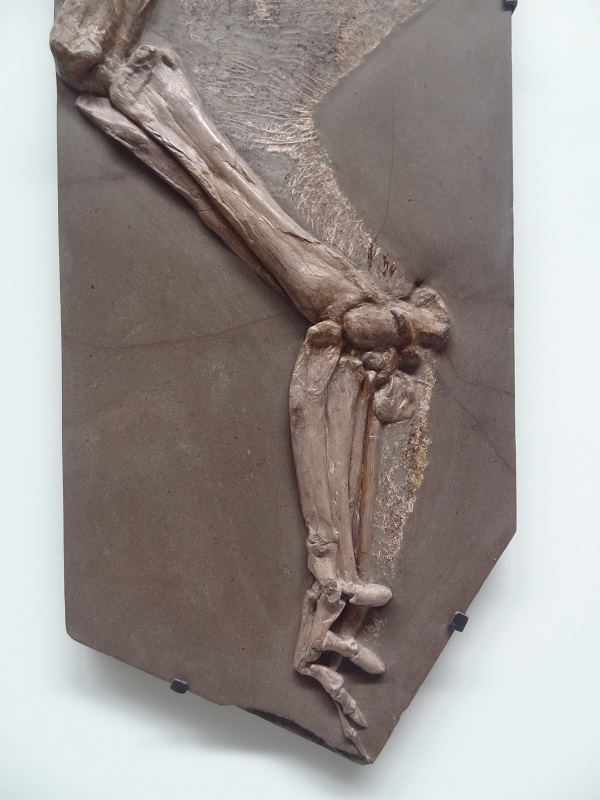










































{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}