

Basilosaurus was just big in the news when a new study about its feeding habits and the predation on young Dorudons was published. This is probably a good time to feature a new reconstruction of the „king lizard“ that I just finished. Originally this was not even made to jump on the Basilosaurus bandwagon but I was looking for an animal which I could use for a certain little art project, and it just fitted perfectly, mainly due to its extreme body proportions. But it became quickly more and more complex and the initial little illustration evolved into much more. I wanted to make a reconstruction which included the aspects which I already discussed in length in my earlier blogposts about the cranial soft tissue of archaeocetes and the highly likely presence of facial vibrissae in this animals. Now I want to explain in detail how and why I reconstructed my Basilosaurus in the way I did.
The Body

I used the famous skeletal drawing of Basilosaurus isis by Kellog (1936) as main reference, however not the original but a modified version by Cameron McCormick with the updated number of vertebrae based on the examination on the vertebrea counting of three overlapping skeletons (Gingerich et al 1990) and a paper by Zalmout et al. 2000.
{kind=link}

As a result of this, this Basilosaurus became even more elongated than Kellogg’s original drawing indicated. This resulted also in some problems, as the whole body was so strongly elongated that I could hardly work on it on the screen at once. Instead I had to work on sections and resize the view again and again to see how it works. One of the main references for its body shape was the ribcage, which shows us how thick its body was about. As Basilosaurus had an extraordinarily long abdomen, I made it somewhat more voluminous than the chest, similar to what we see in many modern cetaceans. To find the basic shape of the body was the most important thing to start until I went more into detail. I had to change my initial draft considerably when I re-checked the ribcage and forelimbs with the skeleton to get the position if the shoulder girdle in the correct way.
Head and mouth

Not surprisingly, the head took especially much time to illustrate, as it includes the most detailed features. I followed the same considerations as in my earlier cranial reconstruction of Dorudon. The line and corner of the mouth were particularly tricky, and it took a long time and many changes until I was finally happy with it. Many reconstructions of archaeocetes shows them with a very deep corner of the mouth, what gives them a quite reptilian appearance. But this animals were mammals, and had after all still the same basic masticatory musculature as us. This includes the masseter muscles, which were quite well developed in archaeocetes. We can see quite well the original position and shape of this muscles from the zygomatic arches. Furthermore there were some other masticatory and facial muscles as well, which affected the shape and position of the corner of the mouth.
Lips

If you have read my earlier posts about archaeocetes, you will know that I am a strong advocate against the common shrink-wrapped reconstructions of archaeocetes with skull-faced heads. I think there are several reasons why we see this type reconstructions so often. People look at those nasty teeth in the jaws and simply assume that they must have been well visible in the living animal. The vague similarity of the skull proportions with those of crocodylians likely contributes to this as well. For exactly this reason we see nearly every year cases in which the rotten remains of odontocetes are mistaken for „prehistoric“ reptilian sea monsters, like this orca carcass from New Zealand.
{kind=link}
Another important point is that artists usually like drawing teeth, and depicting an animal in a badass way is clearly preferred over a „lame“ look in many cases. I was actually surprised how awesome and badass those formidable Basilosaurus teeth still look in the final reconstruction, even with the upper teeth fully covered by the upper lips. It looks now much lesser than the usual crocodile-faced depiction with bare upper teeth. I think most people don’t realize that a lipless Archaeocete could never close its mouth. Even when its jaws would be fully closed the teeth would be still fully visible and the cavity of the mouth constantly in contact with the surrounding water. We see such a condition only in some few highly specialized modern cetaceans like the Ganges river dolphin or the pygmy and dwarf sperm whales. But nearly all other odontocetes have upper lips and cheeks that cover their upper teeth (except of course oddballs like narwhals, but the still have lips). Even odontocetes with large teeth like orcas and false killer whales have fully lip-covered upper teeth. We can be also quite sure that the most archaic and still amphibious whale ancestors had lips as well – like other artiodactyls and mammals usually do. It would be really weird if archaeocetes reduced and even lost their upper lips and their descendants re-evolved them later again. It is also surely important to keep in mind that the general jaw shape dentition was not even that different between the early amphibious forms and the fully marine archaeocetes like Basilosaurus. So lips surely did work around this arrangement and shape of teeth.
What’s just uncertain is the flexibility of those lips and when archaeocetes had still more flexible and perhaps partially retractable lips similar to seals and when already stiff lips like most modern odontocetes. Perhaps the later forms in-between had a somewhat similar condition as we see in beluga whales. I tried to place it somewhere in between.
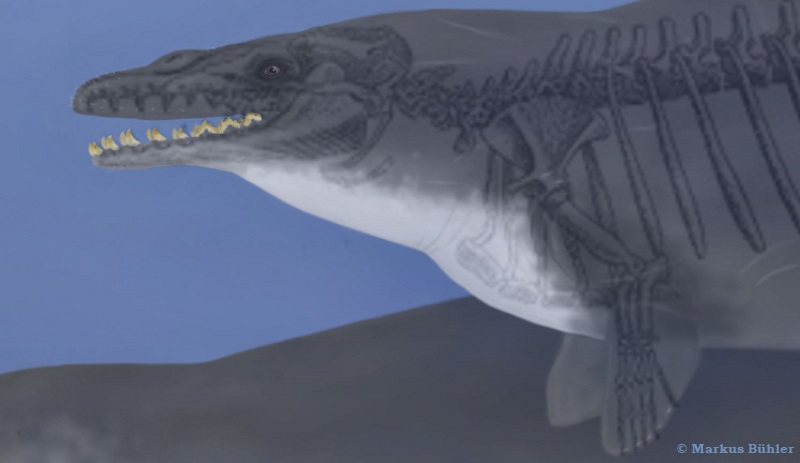
Vibrissae
I also added some vibrissae on the top of the snout and upper lip, at top of the mandible and a few ones behind the nostrils. This are the areas where we still find vibrissae on modern baleen whales and in hippos as well. The whole topic of vibrissae in archaeocetes is quite complex, if you want to read more about it, take a look at this older article.
As a reminder you can see here the vibrissae of a North Atlantic right whale:

Nostrils
Take your fingers and touch your nose. Seriously, do it. Touch it on its back, at the base, the sides and the top. Move it. What do you feel?
You will feel your Os nasale, the different cartilages on the sides and on the tip, and you will also feel the tiny muscles which can use to move the sides of your nose. We humans have somewhat weird looking noses, but they have still the basic mammalian nose structure with a lot of soft tissue over the skeletal opening of the nasal cavity. For some reason comparably many reconstructions of archaeocetes simply follow the outlines of the skull and only show some small openings directly over the nasal cavity.
But where are all the other parts of the nose? In odontocetes, we don’t see much more than the blowhole as all the other parts of their complex and asymmetrical nasal elements are hidden within the rounded melons on the top of their cranium. The only exception is the sperm whale which has a well visible lump of tissue around its blowhole which is – unique in cetaceans – on the left side on the very top of its giant spermaceti organ.

Baleen whales are a much better comparison with archaeocetes here. They lack the melons of odontocetes and have comparably little soft tissue on the top of their skulls. So the dorsal shape of the head is still quite close the the underlying skull – except for the fleshy hump which includes the two blowholes and the accompanying cartilages and muscles.

Neck
The neck of archaeocetes was still longer and much more flexible than those of modern whales. But this still does not mean that the head and torso were as strongly separated as we see it in some reconstructions. I made the neck and throat similar to those of a seal, with a slight indication where the neck ends and the thorax begins. In odontocetes with quite flexible necks like belugas, snubfin dolphins or Amazon river dolphins we see also comparable shapes, which differs from those if other whales with stiffer necks.
{kind=link}
{kind=link}
{kind=link}
Some reconstructions of archaeocetes have quite weird looking skinny necks which strongly separate the head from the body, what´s again not likely at all.
Flippers

The forelimbs of archaeocetes like Basilosaurus were still surprisingly underived compared with the flippers of modern whales and even the forelimbs of seals. They are comparably long and show neither really strong attachments for muscles nor elongations of the fingers. The reason is possibly the fact that archaeocetes had not that much use fore their forelimbs. The basal amphibious forms used their strong hind legs and tail to swim and maneuver underwater, similar to many modern amphibious mammals like otters, beavers, nutrias or even desmans and water opossums. In those animals we find often well developed webbing of the hindfeet, but much lesser on the fore feet.

We see the probably most derived adaptations to a marine habitat of a modern „walking“ mammal in the sea otter, which spends nearly its complete life in the water. This surprisingly large durophagous mustelid has only very short arms, whose main function is to find and collect invertebrate prey, and to crush open hard shells and sea urchins with stones. So I decided to reconstruct the forelimbs not as propper flippers but more like the comparably stumpy and leg-like arms of seacows. I also added a more speculative feature, some vestigial nails. I think so far there was no research to examine the proximal phalanges of archaeocetes to see if there is any indication for such nails. Their superficial structure of the bone could probly solve this mystery, to see if there was still any keratinous growth or not. Seal still have claws, which can be pretty big. Even the highly modified fluke-shaped hind limbs of phocids have short nails. In seacows, which are even fully aquatic we see small hoof-like nails in their forelimbs as well. So even if we don’t have direct evidence for vestigial nails in archaeocetes, we have also so far no direct evidence against it, and the idea really does not seem too far-stretched. So I gave my Basilosaurus seacow-styled protoflippers.
{kind=link}
Tail and fluke

The original version of this Basilosaurus had still a deeper tail and a more conventional shape of its fluke. But I decided to reduce the tail somewhat and make it a bit slimmer, as the dorsal spines of the caudal vertebrae were still comparably short in Basilosaurus. As it also still lacked the caudal peduncle of modern cetaceans I tried to give the caudal fin a somewhat lesser derived shape. The flukes of archaeocetes are really tricky as we can’t reconstruct their shape from the underlying tail vertebrae as in ichthyosaurs and certain mosasaurs or metriorhynchids. Of course we can take a close look at the last caudal vertebrae und see if there’s something going on with them, for example if they are wider and more robust than the vertebrae before them. But this still doesn’t give us really much more information about the original shape of the tail or fluke. Archaeocetes started quite early to use their tails as primary propulsion and their forelegs were likely mainly for maneuverability whereas their hind legs were already fully useless for swimming.
This indicates that a caudal fin was already present in the very early fully marine archaeocetes. But how did it look like? It surely started as a similarly dorso-ventraly flattened tail as those we see in otters and there was also quite likely a stage with a primitive spoon-shaped tail like those of manatees. But the problem is that even with a totally complete line of evolutionary stages between the tail vertebrae of the most basal archaeocetes up to those of modern cetaceans with high dorsal spines, laterally compressed tails and a caudal peduncle for the fluke, we would still don’t know how the caudal fin really looked like.
To get a better understanding of this issue we can take a look at seacows. The manatees which mainly populate freshwater habitats in slow moving rivers or coastal areas (but don’t forget, they also crossed the Atlantic ocean anytime) have only a fleshy oval caudal fin, but it seems to work obviously well enough for them. Dugongs which are living in marine habitats are much more affected by turbulences, waves and similar things and have already a nearly whale-like fluke. They spread over large areas of the globe, over Africa, Asia and Australasia what indicates that they are quite seaworthy. In contrast to manatees they are also much more affected by predation, for example by tiger sharks. In the past they were quite likely also hunted by predatory cetaceans and in some areas possibly also by large marine crocodylians.
This are possibly the reasons why dugongs appear much more whale-like than manatees, as there was a much harder selection for strong swimming abilities than for manatees. Their caudal vertebrae also show elongated dorsal spines and some flattening of the most proximal vertebrae, but we would probably not expect such a well developed fluke from the bones alone. In direct comparison with those of whales it’s however still somewhat lesser derived. The tail is still comparably wide and the fluke itself comparably thick and fleshy.

This could give us some insights into the evolutionary history of whale flukes. I could imagine that there also was a strong selective pressure on the shape of archaeocetes as already little differences of its structure could have had comparably strong hydrodynamic effects. And we also shouldn’t forget that archaeocetes were unlike manatees or dugongs no herbivores but predators which hunted fast and agile prey. That’s why I tend to assume that highly derived oceanic predators like Basilosaurus already had comparably well developed flukes. So I reconstructed the caudal fin as a real fluke and not like manatee paddle, but still different from those of modern cetaceans. It’s still thicker, somewhat longer and more rounded. My very first version was still closer to the flukes of extant cetaceans but I wanted to keep the differences in vertebra shape more in mind and make this also somewhat more obvious in the external shape.
I also gave my Basilosaurus no dorsal fin. One reason for this was because I wanted to make a bit similar to right whale dolphins, little known odontocetes with a very thin and long body, which fully lack a dorsal fin. Another reason was to depict the Basilosaurus as different and more primitive compared with modern whales. However, we have to keep in mind that baleen whales and odontocetes diverged already very early in cetacean evolution, and both do have dorsal fins. So this means that those most basal ancestral baleen whales had either already a dorsal fin (perhaps a very primitive one at least) or that dorsal fins evolved later parallely in both lineages. That´s some food for thought.
Coloration
The coloration of prehistoric animals contains for obvious reasons an especially big amount of speculation, unless we have cave paintings or exceptionally well preserved fossils which can give us some direct evidence for the patterns and colors. In the case of Basilosaurus we have at least some extant relatives, but they can give us still only a vague hint about the colors and patterns of prehistoric whales. I wanted to avoid just another Basilosaurus with the colorations of the Walking with Prehistoric Beasts Basilosaurus. It really doesn’t look that bad and it’s not implausible but I just wanted something different. I’ve discussed the coloration of large marine animals for the paint job of the Meyerasaurus by my friends of kamyk.pl for the museum at Lodz at length here. For the Basilosaurus I also made a first version with a similar color scheme that was somewhat reminiscent to those of a leopard seal. Then I tried a more homogeneous and darker main color which was inspired by false killer whales and long-fin pilot whales, which have also quite elongated bodies. I also made no classical counter-shading with a generally lighter underside. This is more a trend than a rule and a large number of marine tetrapods, especially seals but also a lot of whales, show no real counter-shading at all. Instead I made the area of the throat visually distinctive with a white patch, something we also find in many different modern whales. Because of the incredibly elongated body of Basilosaurus I made also a small white patch in the area of the small hindlimbs. I could imagine that such an eyecatcher of the genital area could make sense in an animal which had possibly really some obstacles to find the right position during mating. We find also similar white eyecatchers (or, as I call them, cetacean arseantlers) in a lot of very different cetaceans, from right whales to sperm whales and various other odontocetes. This brings us to the next point.
{kind=link}
Sexual dimorphism
One of the great things about digital painting is that you can recombine your work again and again. So I made just another Basilosaurus from the first one, what gave me also some more good chances to depict some other aspects. At first I scaled it down to show the sexual size dimorphism. Male Basilosaurs were about 20% longer than females, based on studies of their bones. The second Basilosaurus was also a good opportunity to depict it with a closed mouth. Its mouth is now fully shut and the lips make the teeth no more visible. Remember this couldn’t work at all without upper lips, the teeth would be exposed all the time.

To make the two somewhat more individual I changed the position of the forelimbs of the male to indicate a certain approaching position towards the female. The male has also a slightly lighter coloration, something we also see in older males of many cetaceans like sperm whales or bottlenose whales. He received various scars as well to indicate some intraspecific fights with other males. This is also a quite common thing in various odontocetes from sperm whales to many beaked whales and a lot of dolphins. Last but not least he got a genital organ as well. In fact this was the very first time that I have ever depicted something like this. But prehistoric animals were not always just fighting, hunting and eating and even a gigantic apex predator like Basilosaurus had a social life and sexual interaction. One could even say that next to everything an animal does, no matter if it’s hunting, eating or fighting, has the main purpose to ensure that it can successfully reproduce during its life. So here we go, Basilosaurs having fun.

Background fauna
You shouldn’t underestimate the background fauna. Sometimes it can be somewhat annoying and time-consuming to paint it, but if you have already spent a lot of time on your main subject you can also give it some nice substitutes. You should see the background fauna as a chance and not as a time waster. It gives you a chance to depict some interesting and possibly hardly ever shown species with comparably little effort (unlike a full life-depiction of those animals being in the focus themselves). For this reason you should also avoid totally random stereotypes but really take some time to make a correct depiction. Fish are quite typical elements for background or „prey“ fauna but unless you make a swarm of really tiny and individually unrecognizable fish you should really try to make a realistic depiction. If you are not really familiar with the animal you want to depict, you will very, very easily make errors. I write this because that’s exactly what happened when I started with some unspecified random fish. So I was searching for something better and decided to make a swarm of mackerels. They have a very good fossil record with great specimens from the Eocene which were already very similar to the modern forms. They would also fit within a scenery in the open water and their familiar appearance gives also some more scale for the size of the Basilosaurus. Furthermore I could depict a visually more interesting fish than just one with silver-white colors.

There are also some recycled nautiluses in the background to make the fauna a bit more diverse and to add something that appears somewhat ancient and primordial.
I spent also a considerable amount of time on a Dorudon which was originally only planed as a simple background illustration but at the end it became a similarly detailed reconstruction as the Basilosaurus itself. At first I wanted to depict Basilosaurus hunting a juvenile Dorudon, but then I wanted to depict it next to an adult specimen. The skull length of Basilosaurus and Dorudon is surprisingly similar, despite the very different total lengths. Those of Dorudon has however more elongated jaws and is lesser massive than those of Basilosaurus. But this was still a pretty big animal, about as large as a false killer whale, and with really large jaws.

The Dorudon in detail:

What I really want to emphasize here is that I don´t think this particular reconstruciton is the one and only possible. That´s why I wrote where things were more speculative but also why I made it the way I did. Some traits like the upper lips and the vibrissae (perhaps lesser pronounced, but perhaps even more) were nearly certainly present in archaeocetes, whereas other traits like the shape of flippers and fluke or the presence of a dorsal fin are much more of an issue.
References:
Gingerich, P. D., B. H. Smith, and E. L. Simons. 1990. Hind limbs of Eocene Basilosaurus isis: evidence of feet in whales. Science, 249: 154-157.
Zalmout, Iyad S. 2000. Priabonian Basilosaurus isis (Cetacea) from the Wadi Esh-Shallala formation: First Marine Mammal from the Eocene of Jordan. Journal of Vertebrate Paleontology 20(1):201–204.
Please allow me to compliment you on having pulled together so excellent a rendition of ‚Basilosaurus‘ (It never ceases to amuse me that a relation of the earliest whales will be called ‚lizard‘ for as long as palaeontologists insist that seniority trumps clarity when assigning names*); I love the way you’ve been able to evoke Basilosaurus‘ affinity with whales while very clearly showing that this is quite a different sort of beast, not to mention all the little details you have included so meticulously (for the sexual dimorphism to those little snout-whiskers and even the mackerel!).
I am also glad that you are willing to try something different from the WALKING WITH BEASTS colour scheme; while I would love to see that colour scheme applied to your take on the animal’s anatomy, „variety is the spice of life“ I am also happy to see such a skilled creator furnish an old favourite with such a fine new look!
*Zeuglodon cetoides is definitely a more accurate description, but came along too late!
Keep Well and please keep up the Good Work. (-:
Thank you very much! I will also soon publish another blog article which deals with color schemes of prehistoric marine tetrapods, similar to the one about the Meyerasaurus.
Huzzah! (-:
I shall continue to keep an eye on this website and look forward to seeing your next article!