The fauna of the area which would later become the Messel pit was quite rich in crocs. Some of them strongly resembled modern crocodylians, but some of them were oddballs which differed strongly from any extant species.
The big reptile showcase of the Messel exhibition in the Senckenberg Museum at Frankfurt
The biggest crocodylian of Messsel was Asiatosuchus germanicus, a species which is known from skulls (including the mandible length) up to about 68 cm in length. The total length was up to about 3,5-4 m, about the size of an average American alligator. The very broad and robust jaws also resemble those of an alligator, and it is likely that Asiatosuchus had a similar ecology.
Asiatosuchus germanicus skull, Senckenberg Museum Frankfurt
Asiatosuchus was also the largest animal found in the Messel pit and also the largest carnivore. In contrast to most of the fossils from the lake sediments which had to be transfered from the surounding matrix into an expoy-layer during a complex process, some of the larger croc fossils were massive enough to prep the bones completely out of the oil-shale.
Asiatosuchus germanicus skull, Senckenberg Museum Frankfurt
Here is the partially preserved postcranial skeleton:
Asiatosuchus germanicus hindquarters, Senckenberg Museum Frankfurt
The other Messel crocs were considerably smaller. Diplocynodon darwini for example, an alligatoroid, reached only lengths of about 1,5 m.
Diplocynodon darwini fossil, Senckenberg Museum Frankfurt
This species is knwon from many very well preserved skeletons.
Diplocynodon darwini fossils, Senckenberg Museum Frankfurt
Even tiny hatchlings were preserved.
Juvenile Diplocynodon fossil, Senckenberg Museum Frankfurt
Another small diplocynodontine crocodylians was Diplocynodon deponiaa (formerly Baryphracta deponia). Sadly the only photo I took was rather bad in quality, but at least you can still see one of the most noticeable traits of this species, its very well developed osteoderms:
Diplocynodon deponiae fossil, Senckenberg Museum Frankfurt
Those crocodylians were all comparably similar to some of our modern species. But there were also very distinct forms which have no ecological equivapents today. One of them was the alligatorine Hassiacosuchus haupti (formerly Allognathosuchus haupti). This was a also a rather small animal which only grew to about 1,5 m. It had an extremely shortend snout and wide blunt mushroom-shaped teeth in the posterior part of its jaws. This indicates a certain specialization for small hard prey animals like snails, crabs or small turtles.
Hassiacosuchus haupti fossil
The coolest Messel-croc is sadly only known from very fragmentary remains, a partial mandible and a partial skull (which was sadly not on exhibit) of a single specimen found in the shales. It was Bergisuchus dietrichbergi, a small sebecosuchian. This lineage of crocodyliforms is especially well known from South America, where they evolved a big diversity with some particularly large species. This animals were totally unlike any modern crocodylians and resembled to some degree the large predatory archosaurs from the Triassic like Batrachotomus. They were fully terrestrial and agile and long-legged hunters. Their deep jaws had laterally flattened teeth with sharp serrated edges like those of theropod dinosaurs, but with a pair of enlarge canine-like teeth in the mandible. This animals are very little known and hardly ever mentioned even in paleontology books, but for millions of years those animals were among the dominant terrestrial predators in many parts of the world.
Cast of partial mandible of Bergisuchus dietrichbergi
Bergisuchus was not even the only terrestrial croc of Messel. Another species, originally described as Pristichampsus rollinanti but now dedicated to Boverisuchus is also known from this site, sadly only from extremely fragmentary remains, without any specimens in the exhibition. Here is a speed painting by Joschua Knüppe of Boverisuchus.
Boverisuchus by Joschua Knüppe
This animals had lesser deep jaws than the sebecosuchuians but shared with them the ziphodont meat-slicing dentition. Boverisuchus reached lengths of up to 3 m, but to the usually rather small mammals of the Messel-fauna they must have appeared like huge monsters.
In the next part I will cover the other reptiles from the exhibition.
The Messel pit is one of the most spectacular fossil sites of the world. It gives us an incredible insight into a subtropical ecosystem from the Eocene, about 47 million years ago. The special conditions of this Lagerstätte did not only preserve the bones of the animals which lived in and around a deep crater lake, but also in some cases hair, feathers and body silhouettes, even insects with their colors still preserved. It enables us to get a vivid view of a bygone world, from its plants to insects, fish, amphibians, reptiles, birds and mammals.
It was a weird world which consisted of the survivors of the mass extinction at the end of the Cretaceous 66 million years ago and lineages which had just started to evolve into many animals which we know today. We find a lot of animals which appear surprisingly familiar and modern to us, yet considerably out-of-place in many cases, but also weird archaic lineages and overturned experiments of evolution. In this series I want to feature some of the fossils at the Messel pit exhibition of the Senckenberg Museum at Frankfurt, which I visited in early spring, just before the country was hit by the pandemic. I don´t want to go too much into the details of the species, as this would easily go beyond the scope of this blog article.
I will start the series with the fishes from the exhibition. The most famous fish from Messel pit are quite likely the gars, which I featured already in an earlier blog article several years ago. Today we find gars only in the New World, mainly in the east of the United states, at Cuba and parts of Mesoamerica. But 47 millions years ago gars did swim also in the area which would later become Germany. Gars have interlocking ganoid scales with a very hard layer of enamal on the surface and often fossilize particularly well. The most common species of gar from Messel was Atractosteus strausi (Syn.: A straussi, A. straußi, M. messelensis and A. kinkelini):
Atractosteus strausi, Senckenberg Museum Frankfurt, photo by Markus Bühler
This is a quite common fossil of Messel pit, and you can find similar specimens in many museums around the world. It looked quite similar to modern gars, but remained much smaller than any of the extant species, usually only 20-30 cm on average and about 40 cm at maximum. This is really tiny compared to the giant alligator gar which can reach record lengths of about 3 m. Gars are quite archaic fish which differ from most other extant bony fish in a number of anatomical traits, like their ganoid scales or the heterocercal caudal fins.
Atractosteus strausi, Senckenberg Museum Frankfurt, photo by Markus Bühler
Here is another fossil, fully prepped out of the surrounding matrix:
Atractosteus strausi, Senckenberg Museum Frankfurt, photo by Markus Bühler
Besides A. strausi we find also another small, yet considerably weirder gar, Masillosteus kelleri. It was somewhat bigger than A. strausi and differed from all extant gars in its short and robust jaws which were not adapted to feed on fish but to crush hard-shelled invertebrates with its big and flattened teeth. Sadly there were no fossils of this species on display, but you can see a great life reconstruction of A. strausi and M. kelleri by Joschua Knüppe here:
Fossil gars of Messel pit by Joschua Knüppe
Besides the gars was also another primeval predator, the amiid Cyclurus keheri:
Cyclurus keheri fossil, Senckenberg Museum, Frankfurt. Photo by Markus Bühler
Amiids are a very ancient lineage of bony fish and are not very closely related with any other modern fish. Like the gars they have a heterocercal caudal fin. It was the large fish of this waters and large specimens reached lengths of around 70 cm.
Cyclurus keheri fossil, Senckenberg Museum, Frankfurt. Photo by Markus Bühler
In the Mesozioc there was a huge diversty of amiids, but today there is only one single remaining species, the bowfin (Amia calva) which is restricted to the North American continent. Besides its unusual caudal fin it doesn´t really look particular primitive, but it´s nearly some kind of freshwater analogue of the famous coelacanths of the genus Latimeria. Cyclurus keheri was quite similar to the modern bowfin in appearance, perhaps a little bit stockier. Here is a photo of a modern member of Amia calva by Arthur Kosakowski for comparison:
Modern bowfin (Amia calva), photo by Arthur Kosakowski
Take a close look at the fossil and its well developed teeth, which are very similar to those of the modern bowfin. They aren´t visible in the living fish, because they are hidden by the fleshy lips. Without living relatives for comparisons, we would quite likely only see quite toothy life reconstructions of it. Here is also another nice photo of a juvenile bowfin by Kurtis Smith:
Juvenile bowfin (Amia calva), photo by Kurtis Smith
We know similarly sized specimens from C. keheri from Messel, and tiny gars as well:
Fossil eel, juvenile bowfin, Thaumaturus intermedius and juvenile gars. Senckenberg Museum, Frankfurt. Photo by Markus Bühler
The small fish below the bowfin is Thamaturus intermedius. Its taxonomical relations are still somewhat problematic, some of the sources I found say it was related with mooneyes, whereas other sources say it was a member of the Salmoniformes. There are only two remaining species of mooneyes today, which also live solely on the North American continent today, but have a good fossil record from other continents as well. Despite their unremarkable appearance they belong to a quite archaic lineage and have some quite interesting relationships with the Osteoglossiformes which include such remarkable species like arapaimas or arowanas.
On the left you can see a fossil of the eel Anguila ignota, which is the rarest of all fishes from Messel pit, known only from a single specimen.
The ichthyofauna of Messel pit is quite low in species number and appears surprisingly unexotic, unlike the oddballs which we find among the rest of the contemporary fauna (more about them in the next parts of the series). What I find really interesting is that it includes so many lineages which are now restricted the New World, North America in particular.
Another predator was Amphiperca multiformis, a small but particularly large-headed and big-mouthed perch with a deep body, which looked a bit like a small and shortened version of a warmouth (Lepomis gulosus).
Fossils of Amphiperca multiformis, Senckenberg Museum, Frankfurt. Photo by Markus Bühler
I made a simple speed-painting based on a skeletal drawing of Amphiperca. I usually spend much more time to make fine details and such things, but this time I really only wanted a quick illustration which was done in little more than an hour. I used some color morphs of the highly variable warmouth as main color reference. The fins of this fish are folded, because it´s directly based on the fossil of a dead fish. When the fish was still alive they were of course flexed and looked slightly different, but I wanted to stay as close as possible to the actual proportions of the fossil, so I didn´t try to reconstruct them in a different shape.
Speed-painting reconstruction of Amphiperca multiformis by Markus Bühler
There is also another small perch known from Messel pit, Palaeoperca proxima, whose fossils are however not as common as those of Amphiperca. It was about 20 cm in total length and had a more elongated body than Amphiperca and the two dorsal fins were distinctly separated. Its proportionally much smaller mouth indicats that it also mainly feed on comparably small prey. Sadly there were no fossils of this species on display. The next part of the Messel pit series will cover the reptiles.
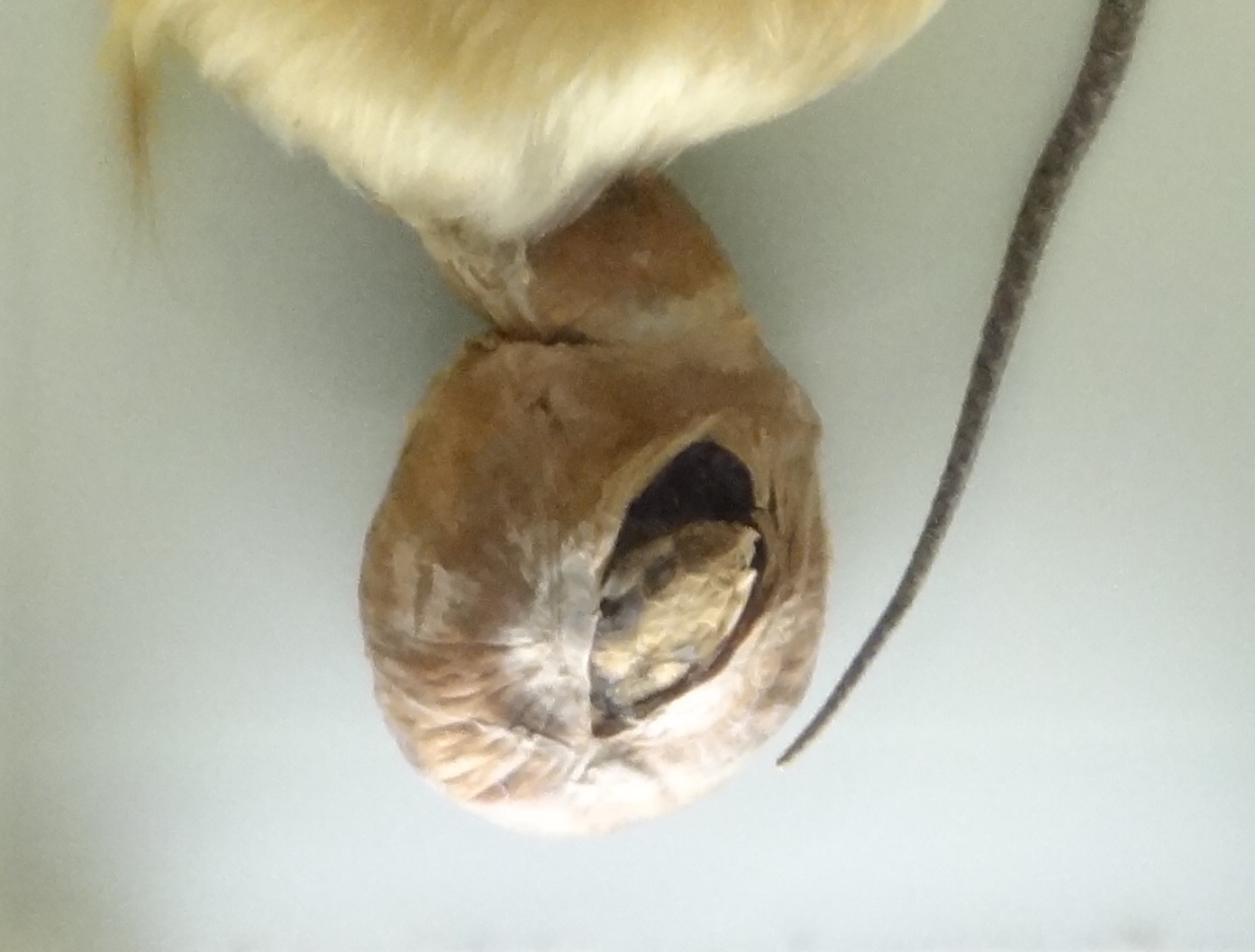
Bizarre deaths and accidents of animals were already covered for several times on the blog, especially in the shit happens-series. Today I want to feature an interesting specimen to continue this topic. This is a partially dissected great crested grebe (Podiceps crestatus) which was found dead, after it tried to gobble an adult European grass snake (Natrix natrix).
Great crested grebe which suffocated while trying to swallow a European grass snake. From the collection of the Senckerberg Museum at Frankfurt. Photo by Markus Bühler
I appears that the snake´s body somehow formed a knot around the grebe´s lower jaw, with fatal effects for both opponents. Great crested grebes usually prefer much smaller prey items like small fish in the range of 10-15 cm, sometimes also amphibians and invertebrates. The normal maximum length of their prey is about 25 cm, so this grebe was particularly enthusiastic when it trid to swallow a whole grass snake which likely exceeded its own body length. You can even see the snake´s head in the opened stomach:
Head of the grass snake in the opened stomach cavity of the grebe
The snake inside the grebe
To give you a better idea of the size of the snake inside, I´ve tried to make a simple reconstruction of it. I have to emphasize this is really just a speculative reconstruction, without an X-ray image it´s hard to say how exactly the snake was curled inside the stomach and perhaps the curve I expected for the snake´s body inside the grebe´s esophagus was not curved enough. But anyway, here it is:
Reconstructed grass snake inside the grebe
The tragic death of the grebe and the snake aside, we see here also an interesting example how unpredictable animal behavior can be. Who would have expected that a grebe would try to attack a snake which is several times as long as its usual maximum prey size for example?
The dissected grebe sadly doesn´t really show the beauty of this elegant kind of waterfowl, so I wanted to finish this blog entry with a somewhat lesser macabre photo. I see grebes comparably often in the wild, but they are usually too shy and too far away to take good photos of them. But several years ago I managed to take some better close-up photos of some grebes at Lake Garda.
Great crested grebe, Lake Garda. Photo by Markus Bühler
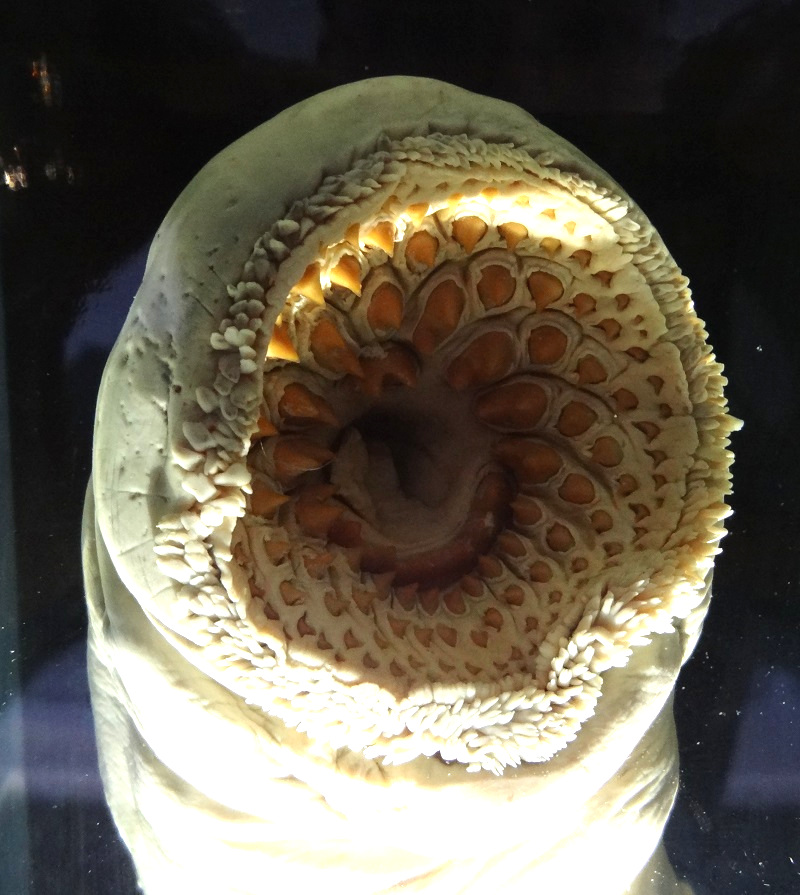
I haven´t posted new stuff on the blog since quite some time, but not because I´m no more writing, but due to the amount of work which is going into some future blog articles. For that reason I was looking for something interesting I could post until the next bigger article is finished. I found a really interesting photo of the mouth of a sea lamprey (Petromyzon marinus) from the Zoological Museum at Kiel at my archive. It looks so wonderfully monstrous yet disturbingly aesthetic for its sheer obvious efficiency that I just had to share this:
Mouth of a sea lamprey (Petromyzon marinus), Zoological Museum Kiel
Sea lampreys are the largest lampreys and can reach lengths of over 1 meter, but usually stay within the range of 70-90 cm. The adult sea lampreys make living as parasites of larger fish, which they attack with their sucker-like mouths and keratinous pseudoteeth and tongue. They rasp and cut pieces out of the fish´s skin until they reach the underlying tissue, so they can suck the effluenting blood, lymph and shreds of tissue.
They became especially infamous after they invaded the Great Lakes and caused havoc among the populations of local fish like lake trouts (Salvelinus namaycush). Here is a taxidermy cast of a namaycush with an attached sea lamprey from Redpath Museum, Montreal:
Sea lamprey attacking a lake trout, Redpath Museum, Montreal.
Many animals have developed amazing anti-predator adaptions, from mimicry to the ability to autotomize certain parts of their own bodies. One of the most bizarre defense mechanisms among mammals is found in the two exant members of the genus Kogia, the pygmy sperm whale (Kogia breviceps) and the dwarf sperm whale (Kogia sima). This dolphin-sized relatives of the enormous sperm whale (Physeter macrocephalus) evolved a method to confuse predators which is…well… special.
Both pygmy and dwarf sperm whales are comparably slow and often bask motionless near the surface. This makes them an easy prey for predators like great white sharks or orcas, which were already found with Kogia remains in their stomaches. They were rarely and just very locally targeted by human hunting activities, for example off Japan, but their passive behavior on the surface made them also an easy target for whalers who could easily harpoon them from nearby boats. When harpooned, they were observed to emit a reddish-brown fluid from their anus which formed a cloud in the water. For this reason they are called Tsunabi in Japanese, a name that can be translated to something like „firecracker-whale“. Sometimes this fluid also leaks out of the anus of stranded specimens, what lead to their Sri Lankan name lie mulla, what means „blood dolphin“. The cloud formed by the dark fluid can apparantly be up to about 100 square meters in size.
This fluid is stored within an a unique sac-like bulging of the lower intestines which can hold up to about 12 l in large specimens of K. breviceps. The consistency of this reddish-brown fluid was compared with chocolate syrup (keep this in mind if you eat chocolate syrup for the next time…), but it can be also have a more granulous sand-like structure that dissolves in the water. Despite its location within the lower intestines it doesn´t seem to be just of fecal origin. It was already found in a foetal K. breviceps and a new-born and still un-nursed K. breviceps, what indicates that this substance which contains large amounts of carbon (up to more than 60%) is not just a digestion-by-product of their cephalopod-rich diet but is synthesized in the guts.
In one case a female K. sima and her calf were together with several dolphins accidentally caught in a net for tunas in the eastern tropical Pacific. The dwarf sperm whale emitted dark reddish fluid and tried to hide in the cloud whenever one of the dolphins approched it or the calf. This happened for several times, what also means that they usually don´t realease the full storage of intestinal fluid at once. This mechanisms, which appear to work in similar way as the inking of cephalopods, irritates the attackers optical and sensorical senses. This would also work in the dark waters at depths of 100 m and more where K. breviceps and K. sima usually hunt, because the olphactorical irritation would still work against sharks. I am not aware of any other depictions of the inking of Kogia, what also encouraged me to create an illustration of this bizarre behavior. Possibly it´s even the first published depiction, but I recently learnt that my good friend Julius Csotonyi also created somewhat earlier an illustration of this behavior for an upcoming book about cetaceans.
Here is also a video which shows a dwarf sperm whale which was chased by a fur seal at Hout Bay Harbour, Cape Town. The dwarf sperm whale released its gut fluid, which immediately produced a huge cloud within the surrounding water:
The reason why kogiids evolved this unusual defense mechanism is possibly linked to heavy predation, which could have an even more severe impact on their populations than we know from direct evidence. Pygmy and dwarf sperm whales have short life cycles and practice some sort of live-fast-die-young-way of life. They are sexually mature at 2,5-5 years, females have (unlike Physeter) a comparably large number of offspring during their lives and they ususally don´t live particularly long. This could also explain why they evolved such a special way to compensate their low speed and agility as a mean to escape from predators. Surprisingly, a similar behavior of releasing a cloud of „fluid“ was also recorded in Physeter as a response of stress, like approaching whalers or orcas. As they lack the sac-like bulge of Kogia to store a fluid, it´s likey that they really just release their feces however. This also indicates that this behavior have evolved already at comparably basal sperm whales.
Sources:
Ellis, Richard (2011). The Great Sperm Whale: A Natural History of the Ocean’s Most Magnificent and Mysterious Creature. Zoology. 179. USA: University Press of Kansas.
Plön S. The status and natural history of pygmy (Kogia breviceps) and dwarf (K. sima) sperm whales off southern Africa, PhD thesis. Grahamstown: Rhodes University; 2004.
Willis, P. M., and R. W. Baird. 1998. Status of the dwarf sperm whale, Kogia simus, with special reference to Canada. Can. Field-Nat. 112:114–125.
The narwhal Monodon monoceros is almost a real-life fantasy creature. But not only due to its historical connection with the legendary unicorn – after all, a whale with an enormous tusk growing out of its head is much more fabulous than most legendary animals from ancient bestiaries which don’t even exist. If we would know narwhals only from fossils, we would surely deeply regret that we could never see this marvelous beast alive. But this creature – one of the most bizarre cetaceans which ever evolved – is an extant species and a good reminder that we still live alongside many incredible animals which can easily compete with the most extraordinary beasts of prehistoric times. There is a lot to say about narwhals, and it would be probably easy to write even a whole book only about their iconic tusks. I recently wrote about their rarely shown vestigial right tusks and a bizarre hybrid between a female narwhal and a male beluga.
Much about the exact function and use of the large torqued tusks still remains an enigma. Many assumptions about the functional use were based on examinations of the anatomy of the tusks, their surface, abrasions and wear patterns. Actual observations of active tusk usage are still extremely rare. A recent video shows that it has apparently really an assisting function for hunting however. But as usually only males possess fully developed tusks, it seems obvious that it has a role for sexual competition as well. Broken tusk tips found embedded into the skull bones of other males indicate that narwhals use their enormous teeth for physical interactions with other males as well.
One of the weirdest phenomena you can find within the cetacean literature are cases of broken narwhal tusk tips inside broken narwhal tusk tips. Errmmm…. What!? To avoid further confusion, I will show you some examples from which I took photos at the University of Copenhagen Zoological Museum.
University of Copenhagen Zoological Museum. Photo Markus Bühler
There are three of such tusks on display, however one of them is just a short fragment.
University of Copenhagen Zoological Museum. Photo Markus Bühler
There is a considerable part of the tips broken off, and you can see the cavity which was originally filled by the pulp. And within this cavity is a plug of torqued ivory, which perfectly fills the opened tusk.
University of Copenhagen Zoological Museum. Photo Markus Bühler
There are also longitudinal fractures and some bending of the tusk around the plug, and it looks like it was forcefully pierced into the broken end.
University of Copenhagen Zoological Museum. Photo Markus Bühler
So what happened here? Was there a narwhal with an already broken tusk tip seeking help by Dr. Monodon who used its own tusk in an altruistic attempt to fill the aching fracture with its own tusk and break the tip off? Or to stay more seriously, could it be that it happened by accident that a male narwhal rammed its thin tusk tip during an interaction in the already fractured tusk of an opponent? Or could it be that narwhal bulls touch each others tusks and one tip became by chance plugged into the opening of the fractured tusk?
University of Copenhagen Zoological Museum. Photo Markus Bühler
It really appears that something like this was happening, and the very existence of not just one but three such cases in a single museum could indicate some never observed behavior. However, as interesting as the idea of narwhal bulls accidentally practicing some sort of endodontic treatment to each others tusks appears, those broken tusks have a quite different background. It started with a tusk fracture, perhaps when a narwhal hunted fish on the sea bottom and had a traumatic collision with a rock. The tip of its tusk broke off, leading the whale quite likely in severe agony for some time. If something similar happens to a human, for example when an incisor is fractured and the pulp becomes exposed, it is just a matter of time until the pulpal tissue becomes infected by bacteria and necrotizes, at least if it is not properly treated. Under totally aseptic conditions – something only possible under laboratory conditions with totally germ-free mice or rats in an isolated environment – the exposed pulpal tissue can produce again new dentine at the fracture and seal the wound.
Fractured narwhal tusks can heal in a similar way without the usual necrotizing of the whole pulp and the resulting fatal conditions for the tusk. Perhaps this is due to the extreme size of the pulp and the many blood vessels within that lead to a better exchange of immune cells and lead also to a faster regeneration of the ruptured tissue. The seawater which always surround the wound might help as well. But admittedly, this is nothing but my personal speculation. It’s noteworthy that exposures of pulps in orcas – something quite common in captive specimens and those which specialize in hunting sharks and skates with highly abrasive skin – lead to necrosis of the pulps as well.
But narwhals can heal such fractures. When the cells at the outer area of the pulp-wound manage to produce a new formation of dentine the fracture is sealed. But this layer is apparently not tightly attached to the inner walls of the tusk. For this reason the ongoing apposition of reparative dentine forms over time a twisted plug which is pressed out of the pulp cavity, forming over time a tiny new tusk. And this is what appears to be the broken tip of a tusk rammed into the open pulp. The crack and bending of the fractured end were possibly also a postmortal artifact when the tusk dried.
I found the information about the reparative dentine of the broken narwhal tusks in the 2003 edition of Walker´s Marine Mammals of the World by Ronald M. Nowak. I was sadly not able to track down the original sources of Reeves, R.R., and S. Tracey.1980. Monodon monocers. Mammalian Species, no. 127, 7 pp and Newman, M.A. 1978. Narwhal. which was cited in Haley, D., ed. 1978. Marine mammals of eastern North Pacific and Arctic waters. Pacific Search Press, Seattle, 138-44. If anyone reading this has access to this original sources or any other information about this topic, I would be quite interested.
Technically, all modern birds are dinosaurs, descendents of a lineage of theropods that evolved complex feathers and the ability to fly. But to be honest… most of them don´t really look very „saurian“ anymore. Those fancy feathers, toothless beaks and stumpy tails – usually coupled with a small overall size – just make it sometimes hard to accept their family background. Even if you know that filaments and feathers were in fact a quite common thing among dinosaurs, even if you are well aware of the fact that a lot of dinosaurs were of quite small body size and even if you are quite familiar with the evolutionary origins of modern birds you can still struggle to accept birds as real living dinosaurs. But sometimes you get a reminder about their theropod heritage, and a glimpse into a time when their ancestors were still not trying to conquer the airspace. For example if you take a close look at one of the largest extant birds, the emu Dromaius novaehollandiae. Don´t look at its pretty blue head or the shaggy double-feathers. Just look down at its staggering feet.
The foot of an emu. Photo by Markus Bühler
There is probably really nothing closer to a non avian theropod foot in the modern world. This is how people even today usually imagine a classical theropod foot. If you forget the rest of the body, you can really easily imagine that it belongs to something much more primordial, somethat that had still jaws lined with teeth, arms with big claws and a long tail. Of course there are some anatomical differences, for example the missing hallux of the emu. But it is still pretty awesome that something still walks the earth on such feet.
The ostrich has even bigger feet, but they are unique and utterly bizarre by nearly every standard, the feet of the cassowary are also pretty cool, but with their elongated claws still somehow too non-standard and not really that similar to any dinosaur feet. It it really frustrating that we missed some of the most spectacular birds that ever lived – the giant moas of New Zealand and the elephant birds of Madagascar – for just a few centuries. The feet of this giant birds were quite likely even closer to the feet of a medium-sized non-avian theropod than anything alive today.
I wrote some time ago about the incredibly polymorphous scales on the feet and legs of the ostrich. If we look at the feet of the emu, we can also see some quite interesting anatomical features as well.
Some of the scales on the backside of the tarsometatarsus form massive conical structures. I have to admit that I have no idea what function they have. But those cone-scales are surely something nobody would expect to have ever existed by looking at the bare bones of a fossil. I can´t help, but they could look pretty cool on the legs of a non-avian theropod as well.
The conical scales on the backside of an emu leg. Photo Markus Bühler
Should I ever get my hands on the foot of an emu, I will try to make a similar listing of its scale types and scale arrangements as I did for the ostrich. I can´t emphasize enough how important and helpful it is to look at the anatomy of living animals to reconstruct species which we know only from their bare bones. You will discover a lot of things which you possibly never noticed before, like in the case of the ostrich and emu those weird scale shapes, the volumous pads on the downside of the toes or the way in which the nails abrade. It will also remind you that we are still living in a world full of amazing animals, and that even many familiar species are much more fascinating and unusual than most people think.
I recently visited the Royal Ontario Museum at Toronto, a monumental museum about zoology, paleontology, history, archeology and ethnology. Among the many wonderful objects in the exhibition about ancient Egypt, one artifact caught my eye in particular. It was a very large fishing hook with a massive leader of thick wire elements – all made of bronze.
Ancient Egyptian bronze hook with chain leader. Royal Ontario Museum, Toronto. Photo Markus Bühler
Here is another photo which shows the hook with some large bronze knifes and baskets to give you a better idea about its size. It was about the size of a hook for big-game-fishing in the sea, or for the largest freshwater fishes like big sturgeons or giant catfish. I have to admit I have a penchant for old and exotic fishing gear, and this one is really one of the coolest ones I have ever seen. From a technical aspect I really admire the work of the ancient smith who made it, the well thought-out construction which helped to avoid a twisting of the line which made it also possible to connect the comparably thin hook with a very strong line. The twisted eyes of the hook and the chain elements provided a maximum of durability. The shape of the hook is not just an U like the much simpler hook on the left, but is highly similar to modern hooks. The diameter in the middle of the curvature is especially thick and slightly flattened to withstand heavy loading. Whoever forged this some thousand years ago – he knew exactly what it needs to catch a monster fish.
Ancient Egyptian bronze hook with chain leader. Royal Ontario Museum, Toronto. Photo Markus Bühler
Today we find nearly identical constructions for shark hooks in which a massive swivel protects the line when the shark is spinning, and a short chain against the sharp shark teeth. Here is an example from the Deutsches Jagd-und Fischerei-Museum (Museum für hunting and fishing) at Munich, from a temporary special exhibition about sharks:
Shark hook with swivel, Deutsches Jagd-und Fischerei-Museum Munich
You can see an even more similar example of a historical shark hook which includes also a chain here.
The African continent is particularly poor in big freshwater fish, and only few species grow to sizes bigger than a meter. There is however one big exception, a fish which reaches enormous dimensions, the nile perch Lates niloticus. This behemoth can exceed lengths of 1,8 m and weights well over 100 kg, making it one of the heaviest freshwater fishes of the world. As usual there are a lot of anecdotes and hardly confirmable dates about much bigger specimens, what makes it quite hard to give more precise sizes and weights.
Nile perch from Lake Victoria, Uganda. Photo by smudger888. Note that this photo already includes a certain amount of forced perspective, and in direct comparison with the man behind it, the fish would appear arond 20-25% smaller.
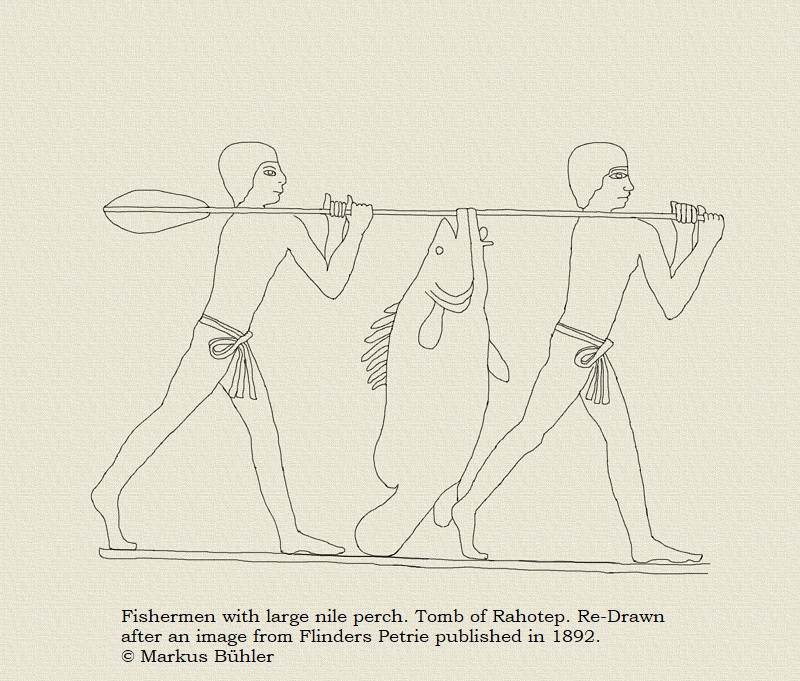
Even before this species was introduced to many bodies of water in which it was originally not native (with desastrous effects, like in Lake Victoria) it had a very large range, which included the nile in North Africa. The ancient Egyptians were well familiar with this fish, and there are even cases known of mummified nile perches, like this specimen in the Egyptian Museum at Cairo. Fish was caught with nets, harpoons and also hook and line (which included simple rods but also handlines), and it was an important source of protein. Most of those fish were however comparably small, species like cyprinids, tilapias or smaller and medium sized catfishes. Catching the really big nile perches was probably not that easy as it required special gear. The fabrication of this hook was probably already a lot of work and bronze was still very valuable even at small amounts at that time.
Fish relief, about 2.500 years BC, Kunst-und Gewerbemuseum Hamburg
Bones of large nile perches were also found among other animal relics at archeological sites, sometimes in form of waste, sometimes as burial objects. Some of them include relics of really huge specimens, like several nile perches from the necropolis of Elephantine which were about 1,8 m in life. One particular specimen was even about 1,9 m. There were also a lot of nile perch bones found at the temple of Satis, which included several very large specimens, including one of about 1,9 m. It´s noteworthy that this is still not bigger than the largest modern day nile perches from the big lakes in their more southern range. Today such huge specimens are no more found at Egypt anymore, but it seems that they were considerably more common in the past, and the ancient fisherman obviously managed to catch them.
As nile perches usually prefer comparably deep water, they were likely rarely caught with harpoons, and it seems likely that they were also no common targets for neting, at least not the big ones. So angling with hook and line in big game style was likely the most efficient way to target particularly big specimens, which had also surely a considerable worth. The ancient Egyptians called the nile perch „the fighter“, what also possibly indicates its behavior when it was caught with hook and line. Those fishermen had nothing even remotely comparably to the modern fibreglas rods and reels, but just handlines to catch those enormous fish which could weigh considerably more than an average human.
If you want to learn more about fishes and fishing in ancient Egyptian art and culture, take a look at those links: 1, 2, 3.
Sources:
BOESSNECK, J. und von den DRIESCH, A. (1982): Studien an subfossilen Tierknochen aus Ägypten. – Münchener Ägyptol. Studien, 40:58, München-Berlin.
I wrote some time ago about zoological treasures in archeological, ethnological and historical museums. Today I want to share some quite interesting examples of such „treasures“ which I have seen in the Oceania exhibition of the Ethnological Museum of Berlin. What you see here is a boar’s mandible from Vanuatu, a group of islands located in the South Pacific Ocean east of Australia.
Boar mandible with circle tusks from Ambae, Vanuatu. Ethnological Museum of Berlin, photo Markus Bühler
The obviously most striking feature of the mandible are the extremely long circular tusks. They are so long that their tips have grown into the lower jaw and penetrated the bone.
Boar mandible with ingrown lower tusks. Ethnological Museum of Berlin. Photo Markus Bühler
You can see how how the infection caused by the tusk tips lead to a degradation of the surrounding bone and a thickening of the bone behind. The ingrown tusks acts like foreign bodies and cause a constant local infection. In the living animal they must have pierced the cheeks as well.
Boar mandible with ingrown lower tusks. Ethnological Museum of Berlin. Photo Markus Bühler
This is an extracted lower boar tusk, which shows how the tooth formed a full circle, with the tip just next to its own apical end.
Extracted circle tusk. Ethnological Museum of Berlin. Photo Markus Bühler
Here is another example, but this boar luckily died before the tusks could impale its jaw.
Boar mandible with still not fully formed circle tusks. Ethnological Museum of Berlin. Photo Markus Bühler
For a considerable time I was wondering how this circle tusks could grow to such grotesque shapes. It was obvious that it resulted from a lack of abrasion with the upper tusk, which usually limit the lengths of the lower tusks while forming constantly sharp edges. Sometimes we see similar cases as a result of misplaced teeth which lead to a missing abrasion between upper and lower tusks. I covered such a case of a pot-bellied pig which had highly elongated lower tusks. The extremely shortened skull and jaws of pot-bellied pigs often result into various tooth displacements. But those jaws from Vanuatu belonged to pigs which were still quite primitive compared with most modern breeds, with long heads similar to wild boars and with well grown teeth.
So how could they grow this bizarre circle tusks? Were those pigs possibly selected for dental misplacement that make such teeth growing“naturally“? The answer is no, those boars were actually results of a surgical procedure which made their tusks growing into this unnatural shapes. Their upper tusks were extracted at early age, so the natural curve of the lower tusks could grow longer and longer without being abraded by an antagonistic upper tusk. The boars selected for this were kept isolated and fed with special food, and apparently they were also castrated. This was likely to avoid accidental fractures of the tusks during fights with other boars and to let them grow better even with the chronic inflammation of the cheeks and jawbone.
But why did Vanuatuans want boars with circle tusks? The answer is pretty simple, because breeding boars with circle tusks was literally like breeding money. Those circle tusks were not just used for decoration or jewelry but as money as well. And they longer they were, the bigger their worth was. They were used to make jewelry like necklaces, as decoration for sculptures and for certain rituals. You have to keep in mind that it takes several years until a boar has grown a circle tusk, and even more for a spiral tusk. During this whole time it needs extra food, and even more than a normal pig, which can also forage in the wild. But when the boar is finally killed and its tusks removed, they are worth a little fortune. I highly recommend to watch this short documentary about the role of circle tusks in modern Vanuatu. You can also see a living boar with circle tusks which pierce its cheeks. There are also a few incredibly extreme spiraling tusks from an ethnological collection featured as well.
You can get an idea about the importance of circle boar tusks if you take a look at the flag and the coat of arms of Vanuatu.
Flag and coat of arms of Vanuatu. Source Wikipedia
The whole museum exhibition at Berlin was moved to another location after my last visit and I don’t know if those specimens will be still exhibited in the new museum. It was the same exhibition which also included the mislabeled Kogia mandible about which I already wrote.
This is mainly an updated version of a blog post which I already wrote in 2009 in German. You can find the original post here.
Kürzliche erregten neue Forschungsergebnisse um einen bizarren Hybriden aus Narwal (Monodon monoceros) und Beluga (Delphinapterus leucas) weltweite Aufmerksamkeit. Er wurde nach Angaben eines grönländischen Inuit namens Jens Larsen 1986 oder 1987 in der Disko-Bucht im Westen Grönlands geschossen, und wies unter anderem einige äußerst ungewöhnliche Besonderheiten der Zähne auf. Der Wal soll grau gefärbt gewesen sein, und seine Brustflossen denen eines Belugas, die Schwanzflosse dagegen eher der eines Narwals geähnelt haben. Larsen beschloss aufgrund des ungewöhnlichen Aussehens des Wals dessen Schädel aufzuheben. Etwa zur gleichen Zeit sollen in der Gegend noch zwei ähnliche Exemplare geschossen worden sein, von denen jedoch eines versank und nicht geborgen werden konnte. Der Kopf des dritten Exemplars wurde zum Skelettieren an der Küste ausgelegt, allerdings niemals wiedergefunden, so dass als einziges physisches Relikt der von Larsen aufbewahrte Schädel verblieb.
Als 1990 der dänische Zoologe und Walexperte Mads Peter Heide-Jørgensen den Schädel auf dem Dach eines Werkzeugschuppens im grönländischen Kitsissuarsuit entdeckte, erkannte er gleich dass es sich dabei um etwas wirklich außergewöhnliches handelte. Larsen stiftete den Schädel für weitere Untersuchungen, woraufhin dieser nach Kopenhagen gebracht wurde. Seine Anatomie zeigte Ähnlichkeiten zu jener von Narwalen und Belugas, wies aber auch eine Reihe von Merkmalen auf, die völlig einzigartig waren, und sich von allen anderen bekannten Walen unterschieden. Anhand der teilweise verwachsenen Suturen (Knochennähte) von Oberkiefer und Zwischenkiefer lies sich erkennen dass es sich um ein erwachsenes und bereits älteres Tier gehandelt hatte. Beim Vergleich mit den Schädeln von Belugas und Narwalen lag der Schädel in seinen Proportion etwa zwischen beiden Arten, in seinen Größendimensionen allerdings etwas über deren durchschnittlicher Größen. Bereits vor einigen Jahren wurde für den Hybriden der Begriff „Narluga“ geprägt, welchen ich hier auch verwenden möchte.
Schädel des Narlugas. Foto Mikkel H. Post, Zoologisches Museum Kopenhagen
Besonders ungewöhnlich waren allerdings die Zähne, welche sich sowohl in Anzahl, Form als auch Größe von jenen von Belugas und Narwalen unterschieden. Belugas besitzen im Ober-und Unterkiefer je 8-11 Zähne auf jeder Seite.
Belugaschädel im Zoologischen Museum Kiel. Foto Markus Bühler
Die Zähne von Belugas sind zapfenförmig, und nutzen sich vor allem im Oberkiefer oft auf typische Weise löffelförmig ab.
Nahansicht der Beluga-Zähne mit den unterschiedlichen Abrasionsformen in Ober-und Unterkiefer. Foto Markus Bühler
Beim Narwal dagegen besitzen üblicherweise nur die Männchen einen einzigen funktionellen Zahn, bei welchem es sich um einen massiv modifizierten linken oberen Eckzahn (er entstammt dem Oberkiefer und nicht dem Zwischenkieferknochen, ist also kein Schneidezahn) handelt, welcher außerhalb des Mauls durch die Haut der Oberlippe wächst.
Narwalschädel im Staatlichen Museum für Naturkunde Stuttgart. Foto Markus Bühler
Normalerweise ist er nur auf einer Seite ausgeprägt, während der deutlich kleinere und auch nicht gedrehte rechte Eckzahn üblicherweise im Kieferknochen verbleibt. Bei weiblichen Narwalen finden sich derartige im Kiefer verbleibenden Eckzähne sowohl rechts als auch links.
Nicht durchgebrochener rechter Eckzahn eines Narwals, Kunstkammer im Landesmuseum Württemberg im Alten Schloss Stuttgart. Foto Markus Bühler
Eine derartig extrem ausgeprägte dentale Asymmetrie wie bei Narwalen ist einzigartig unter den lebenden Säugetieren. Gelegentlich finden sich allerdings auch Exemplare mit einem auch rechts teilweise oder sogar voll ausgebildeten Stoßzahn, oder auch Weibchen mit ein oder sogar zwei Stoßzähnen. Das eigentliche Maulinnere ist dagegen völlig zahnlos. Gelegentlich finden sich auch kleine verkümmerte Zähne an der Basis des Stoßzahns im Knochen des Oberkiefers, welche allerdings nicht durchbrechen.
Der Schädel des Wals aus der Diskobucht hatte dagegen ursprünglich im Oberkiefer auf jeder Seite je 5 und im Unterkiefer je 4 Zähne, von denen allerdings ein oberer und ein unterer postmortal verloren gingen. Aufgrund von Abrasionen und ihrer Position im Knochen kann man davon ausgehen dass sechs oder sieben der zehn Oberkieferzähne zu Lebzeiten in die Mundhöhle ragten.
Ansicht des Oberkiefers von der Gaumenseite mit erkennbaren Bereichen der im Knochen verbliebenen vordersten Zähne. Foto Markus Bühler
Die drei oder vier vordersten Zähne lagen horizontal im Knochen und sind nie in die Mundhöhle durchgebrochen. Ihre sehr langgestreckte gerade Form erinnert etwas an die nicht durchbrechenden Stoßzähne von Narwalen, die für diese typische Verdickung an der Wurzelspitze fehlt allerdings. Die vorderen Zähne ließen sich nicht ohne Beschädigung des umliegenden Knochens entfernen, anhand von Röntgenaufnahmen ließ sich ihre Form und Länge aber ermitteln. Dies ergab eine stattliche Länge von 25,6 cm für den linken ersten Oberkieferzahn und 24,8 cm für den rechten ersten Oberkieferzahn. Die zweiten hatten immerhin noch eine Länge von 19,6 cm und 21,5 cm, während die weiter hinten liegenden immer weiter an Länge abnahmen, dafür aber auch eine mehr oder weniger starke Krümmung aufwiesen. Nach Angaben von Jens Larsen hatte der Narluga welcher im Wasser verloren gegangen war, auch sichtbare vordere Zähne im Oberkiefer.
Einige der aus dem Kiefer entfernten Zähne. Foto Eline Lorenzen, Zoologisches Museum Kopenhagen
Die Zähne im Unterkiefer zeigten ebenfalls eine Reihe von Besonderheiten. Zunächst waren sie ausgesprochen groß und massiv gebaut, deutlich größer als die Zähne von Belugas. Auch der gesamte vordere Bereich des Unterkiefers war äußerst breit und massiv. Aufgrund ihrer Form und Größe müssen die unteren Zähne sogar noch bei geschlossenem Maul teilweise sichtbar gewesen sein. Einige waren merkwürdig verdreht und zeigten vage Andeutungen von Längsrillen, vergleichbar den Stoßzähnen von Narwalen. Die ungleichmäßigen Abrasionsmuster ließen ebenfalls erkennen dass sie sich während des Wachstums leicht um die eigene Achse gedreht haben müssen.
Nahansicht der Zähne im rechten Unterkiefer. Foto Markus Bühler
Die vier rechten und ersten drei linken Unterkieferzähne standen alle recht dicht beieinander, waren von ähnlicher Größe und waren deutlich noch vorne geneigt. Der leider nicht mehr vorhandene vierte linke Zahn saß dabei aber deutlich weiter hinten im Kiefer als sein Pendant auf der rechten Seite, zudem konnte man anhand des leeren Zahnfachs erkennen dass er nicht nur deutlich kleiner gewesen sein muss, sondern auch sichtlich nach hinten geneigt war.
Ansicht der linken Unterkieferhälfte. Man erkennt das leere Zahnfach des deutlich kleineren und weiter hinten liegenden letzten Zahns sowie weiter unten den Austritt des Nervus mentalis aus dem Foramen mentale. Foto Markus Bühler
Als der Schädel erstmals untersucht wurde, war seine genaue Identität noch nicht abschließend geklärt. Ursprünglich wurde auch ein Narwal mit starken dentalen Anomalien für möglich gehalten, die Möglichkeit eines Hybriden allerdings auch schon in Betracht gezogen. Kürzlich vorgenommene Untersuchungen seines Erbgutes durch eine Forschergruppe um Eline Lorenzen vom Zoologischen Museum Kopenhagen konnten nun tatsächlich die Vermutung um seine hybride Herkunft bestätigten. Zudem konnte ermittelt werden dass es sich bei diesem wirklich ungewöhnlichen Exemplar um eine Kreuzung aus einem weiblichen Narwal und einem männlichen Beluga handelte. Die Beschreibung der beiden anderen mutmaßlichen Hybriden durch Jens Larsen ist auch insofern bemerkenswert weil sich hierbei die Frage stellt ob diese alle unabhängig voneinander gezeugt wurden, oder ob sie möglicherweise alle die selbe Mutter oder den selben Vater hatten. Ohne verbliebene körperliche Relikte dieser beiden Exemplare wird sich diese Frage allerdings nie beantworten lassen. In der Diskobucht kommen sowohl Narwale als auch Belugas vor, und sie ist auch eine der wenigen Gegenden in welcher beide Arten auch zu ihrer jeweils unterschiedlichen Paarungszeit anzutreffen sind. Gelegentlich schließen sich einzelne Belugas auch Gruppen von Narwalen an, aber auch einzelne Narwale in Belugaschulen kommen gelegentlich vor, etwa ein junges und möglicherweise verirrtes Narwalmännchen welches sich im kanadischen Sankt-Lorenz-Strom den dort lebenden Belugas angeschlossen hat. Es gibt eine ganze Reihe von dokumentierten Hybriden zwischen verschiedenen Walspezies, sowohl unter Zahnwalen wie Delfinen oder Schweinswalen, als auch von Bartenwalen.
Die Interaktion der unterschiedlichen Erbanlagen ist wirklich erstaunlich. Besonders dahingehend dass jene Gene welche beim Narwal das Größenwachstum und die Torsion des Stoßzahns steuern sich in stärkeren Maße auf die Zähne des Unterkiefers ausgewirkt haben als auf jene im Oberkiefer. Es wäre sehr interessant zu wissen ob, und falls ja wie, sich bei der umgekehrten Kreuzung zwischen einem Narwalmännchen und einem Belugaweibchen die Genkombinationen anders ausgewirkt hätten.
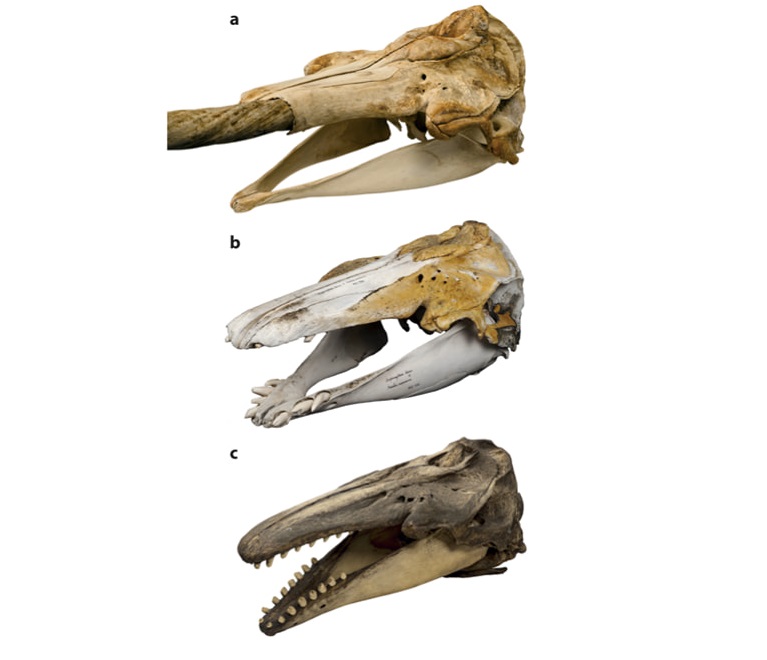
Vergleich der Schädel von Narwal (a), Narluga (b) und Beluga (c). Mikkel Høegh Post, Zoologisches Museum Kopenhagen
Eine weitere erstaunliche Erkenntnis ergab die Untersuchung der Kohlenstoff-und Stickstoffisotpe im Kollagen des Knochens. Demnach ernährte sich der Narluga nicht von den gleichen Beutetieren wie Belugas oder Narwale, sondern erbeutete vor allem Fische in tieferen Wasserschichten. Möglicherweise bevorzugte er aufgrund seiner ungewöhnlichen Bezahnung bestimmte Beute die er nur in tieferen Wasserschichten finden konnte.
Seit ich 2007 zum ersten Mal von diesem bizarren Wal gelesen habe, war ich fasziniert von ihm. Da die ursprüngliche Veröffentlichung von Mads Peter Heide-Jørgensen damals noch nicht im Internet frei verfügbar war, schrieb ich ihn an und bekam freundlicherweise eine digitale Version von ihm zugesendet. Die gescannte Version des Drucks beinhaltete eine ganze Reihe von interessanten Fotos, allerdings leider in sehr unzufriedenstellender Bildqualität. Zudem gab es einige weitere Fotos des Schädels im Internet, viele seiner Details ließen sich aber auch bei ihnen nicht vollends erkennen. Ein Jahrzehnt später, im Frühsommer 2017, bekam ich die Möglichkeit den Narlugaschädel im Original im Archiv des Zoologischen Museums in Kopenhagen zu begutachten, und seine Anatomie vor Ort zu studieren. An dieser Stelle möchte ich mich noch einmal ganz herzlich bei Eline Lorenzen und auch vor allem bei Daniel Klingberg-Johanson dafür bedanken den Schädel und viele andere faszinierende Stücke im nichtöffentlichen Archiv des Museums ansehen zu können.
Schädel des Narluga im Archiv des Zoologischen Museums Kopenhagen. Foto Markus Bühler
Was mich ganz besonders interessierte, war wie dieser Wal wohl zu Lebzeiten ausgesehen haben könnte. Da lediglich der isolierte Schädel, die Verwandtschaftsverhältnisse und die sehr vage Beschreibung von Jens Larsen vorhanden waren, stellte sich hier eine Aufgabe,welche viel eher der Rekonstruktion eines nur von Fossilien bekannten ausgestorbenen Tieres vergleichbar war (etwa meiner Basilosaurus-Rekonstruktion). Bisher gab es keinerlei Bilder welche einen Eindruck vermittelten wie der Hybrid einmal ausgesehen haben könnte (es gab eine einzige Darstellung, welche allerdings lediglich eine digitale Kombination aus einem Narwal und einem Beluga darstellte, und weder die Zähne noch die beschriebene Farbe berücksichtigte), was seine Rekonstruktion umso interessanter machte.
Da das einzigartige Gebiss das hervorstechendste Merkmal des Narlugas war, sollte es bei der Rekonstruktion auch entsprechend gut erkennbar sein. Daher entschloss ich mich direkt ein Bild des komplett artikulierten Schädels aus einer vor längeren Zeit erfolgten Pressemitteilung als Basis zu verwenden. Bei Zahnwalen im Allgemeinen und bei Gründelwalen wie dem Narwal und Beluga im Speziellen wird der Schädel von einer großen Menge Weichgewebe umschlossen. Vor allem die Melone, das Echolotorgan welches oberhalb des Schädels sitzt, nimmt einen sehr großen Raum ein. Dazu kommen noch die Bereiche in denen die komplexen Strukturen zum Blasloch führen, sowie die Lippen und natürlich das dicke Unterhautfettgewebe. Eine sehr hilfreiche Illustration zum besseren Verständnis der Anatomie eines Belugakopfes kann man hier sehen.
Rekonstruktion der Kopfform des Narlugas anhand des Schädels
Bei Belugas kommt noch hinzu dass sie durch Muskelkontraktion die Form ihrer Melone in gewissem Rahmen verändern können, was das Heranziehen von Referenzbildern noch problematischer machte, denn selbst bei ein und demselben Individuum kann auf verschiedenen Bildern die Melone unterschiedlich geformt sein. Belugas besitzen auch für Wale ungewöhnlich gut ausgebildete und bewegliche Lippen. Das alles musste bedacht werden, selbst wenn hier selbstverständlich einiges an künstlerischer Freiheit einfloss. In seiner Kopfform steht der Narluga nun zwischen jenen der beiden Elternarten, mit den eher stärker verrundeten Formen des Narwals aber einer schlankeren Halspartie und etwas voluminöseren Oberlippen um dem Belugaerbe gerecht zu werden. Den fehlenden letzten Zahn auf der linken Unterkieferseite rekonstruierte ich anhand der Größe und Ausrichtung des leeren Zahnfachs im Kiefer. Man sieht nun ganz gut die starke Asymmetrie in der Position und Form der Zähne im Unterkiefer. Bei der Pigmentierung des Maulinneren verwendete ich das Foto eines Narwalmauls als Referenz. Bei Walen findet sich nicht wie sonst bei Säugern üblich eine klare Abgrenzung zwischen dem normalerweise von Schleimhaut ausgekleideten Maulinneren und den von Epidermis bedeckten Lippen.
Narlugakopf mit rekonstruiertem vierten linken Zahn im Unterkiefer
Bei der Körperform stützte ich mich auf die skelettalen Proportionen von Belugas, um anhand des Schädels die dazugehörigen Dimensionen des Körpers festzulegen. Auch hier versuchte ich einen Mittelweg zwischen den zwei Elternarten zu finden. Bei Narwalen ist der Körper sehr stromlinienförmig und glatt, während Belugas einen sehr stark vom Körper abgesetzten Kopf und seitlich am Körper liegende Fettpolster und Längsfalten besitzen. Auch haben sie noch einen wahrnehmbaren, leicht eckigen Rückenkiel, während bei Narwalen die Rückenflosse bis auf eine Linie von kleinen, kaum erkennbaren Hauttuberkeln zurückgebildet ist. Belugas haben größere und deutlich breitere Brustflossen als Narwale, weshalb ich sie aufgrund der Beschreibung Larsens auch als Referenz verwendete. Bei der Schwanzflosse dagegen orientierte ich mich an den stark halbkreisförmig ausgebildeten Flossenlappen des männlichen Narwals. Interessanterweise findet sich bei Narwalen ein Geschlechtsdimorphismus bei der Form der Schwanzflosse, welche allem Anschein nach mit den hydrodynamischen Auswirkungen des Stoßzahns zusammenhängt.
Die Farbe des Narluga wurde als grau beschrieben, und unterscheidet sich insofern deutlich von der weißen, beziehungsweise weiß-schwarz marmorierten Farbe von Belugas und Narwalen. Allerdings sind bei beiden Arten die Jungtiere noch grau gefärbt, und es ist nicht völlig ausgeschlossen dass bei dem Hybriden möglicherweise atavistische Anlagen zum Vorschein kamen, welche aus der Zeit stammten, als ihre Vorfahren noch nicht in polaren Gebieten lebten, und vermutlich auch noch nicht von weißer Grundfarbe waren. Daher bekam der Narluga ein Farbschema welches sich an den Jungtieren der beiden Elternarten orientierte. Im Kopf-und Halsbereich deutete ich eine leichte graue Marmorierung an, welche mehr einem jungen Narwal entspricht, der hintere Körperbereich dagegen der homogeneren Farbe junger Belugas. Zudem nahm ich mir die kreative Freiheit die Brustflossen etwas dunkler vom Körper abzusetzen, wie es in stärkerer Form bei Narwalen der Fall ist.
Narluga mit hypothetischen Farbschema
Um den Hintergrund noch ein bisschen interessanter zu gestalten fügte ich einen kleinen Schwarm Polardorsche (Boreogadus saida) ein, eine Fischart welche auch in der Disko-Bucht lebt, und sowohl vor Flussmündungen bis hinab in Tiefen von 900 m vorkommt. Diese Fische stellen auch eine wichtige Nahrung für Belugas und vor allem Narwale dar.
Polardorsche (Boreogadus saida)
Insgesamt arbeitete ich an der Rekonstruktion mehrere Monate, und trotz mancher Ungewissheiten und künstlerischer Freiheiten bestimmter Details hoffe ich doch sehr dass sie zumindest einen realistischen Eindruck dieses außergewöhnlichen Wals vermitteln kann.
Quellen:
Fontanella, J. E., Fish, F. E., Rybczynski, N., Nweeia, M. T. and Ketten, D. R. (2011). Three-dimensional geometry of the narwhal (Monodon monoceros) flukes in relation to hydrodynamics. Mar. Mamm. Sci. 27, 889-898.
Heide-Jørgensen, M. P. & Reeves, R. R. Description of an anomalous Monodontid skull from west Greenland: A possible hybrid? Mar. Mamm. Sci.9, 258–268 (1993).
Nweeia, M.T., et al. 2009. Considerations of anatomy, morphology, evolution, and function for narwhal dentition. The Anatomical Record 295, 6: 1006-1016.
M. Skovrind et al. Hybridization between two high Arctic cetaceans confirmed by genomic analysis. Scientific Reports. Vol. 9, June 20, 2019. doi:10.1038/s41598-019-44038-0.
Diese Website benutzt Cookies zur Identifikation von Benutzern und zur statistischen Auswertung von Inhaltsaufrufen. Mit der Nutzung erklären Sie sich damit einverstanden.































































{kind=link}
{kind=link}
{kind=link}
{kind=link}